薩爾特里奧獵龍屬
萨尔特里奥猎龙(学名:,意为“萨尔特里奥的猎人”)是角鼻龙类恐龙的一个属,生存于早侏罗世锡内穆阶的意大利,模式种是扎氏萨尔特里奥猎龙(Saltriovenator zanellai)。虽然尚未发现完整骨骼,但一般认为萨尔特里奥猎龙是种大型两足食肉动物,类似角鼻龙。[1]
| 萨尔特里奥猎龙属 化石时期:锡内穆阶早期 ~ | |
|---|---|
 | |
| 模式标本 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 獸腳亞目 Theropoda |
| 演化支: | 新獸腳類 Neotheropoda |
| 演化支: | 鳥吻類 Averostra |
| 演化支: | †角鼻龙类 Ceratosauria |
| 属: | †萨尔特里奥猎龙属 Saltriovenator Dal Sasso et al., 2018 |
| 模式種 | |
| †扎氏薩爾特里奧獵龍 Saltriovenator zanellai Dal Sasso et al., 2018 | |
发现与命名

1996年8月4日,业余古生物学家安杰洛·扎内拉(Angelo Zanella)在意大利北部萨尔特里奥的萨尔诺瓦大理石采石场寻找菊石时,发现第一具萨尔特里奥猎龙遗骸。扎内拉在米兰自然历史博物馆工作,因此该机构在接到通知后派出一个小组去调查该发现。在乔治·特鲁齐的指导下,克里斯蒂亚诺·达尔·沙索和阿尔伯托·卢瓦尔迪成功挖出一些明显含有骨骼的岩块。在出土前不久,这具骨骼被采石场开采大理石的炸药炸成了碎片。石块在固定后被浸入甲酸中1800小时以释放其中的骨骼。[1]最初,报告中说总共收集了119块骨头碎片;[2][3]这个数字后来增加到132。然而,大多数碎片都无法准确识别。[1]
2000年,博物馆举办了一次化石展览。因此,达尔·沙索临时给这具被视为新物种的恐龙骨骼起了个意大利语名字——萨尔特里奥龙(Saltriosauro,有时也被拉丁化为Saltriosaurus),但当时仍为无资格名称。[1]
2018年12月,达尔·沙索(Dal Sasso)、西蒙·马加努科(Simone Maganuco)和安德里亚·考尔(Andrea Cau)将该标本命名并叙述。模式种是扎氏萨尔特里奥猎龙(Saltriovenator zanellai),属名取自化石发现地和拉丁语venator(意为“猎人”,是兽脚类学名中常见的后缀)。作者指出,属名中的后缀venator也是拉丁语中对古罗马角斗士的称呼。种名纪念扎内拉。因为这篇文章是在电子杂志上发表,所以生命科学标识符是使这个名字有效的必要条件。属的标识符为8C9F3B56-F622-4C39-8E8B-C2E890811E74,物种的标识符为BDD366A7-6A9D-4A32-9841-F7273D8CA00B。萨尔特里奥猎龙是第三只来自意大利的恐龙与第一只来自阿尔卑斯山的恐龙,也是继棒爪龙后第二只来自意大利的兽脚类。[1]
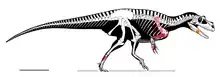
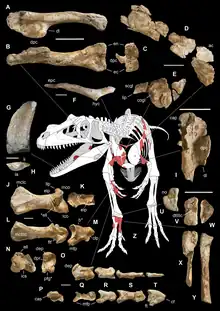
正模标本MSNM V3664发现于萨尔特里奥组,可追溯至距今约1.99亿年的锡内穆阶早期,由下颌碎片组成。已发现约10%的骨骼,包括一颗牙齿、一个右夹骨、一个右前关节骨、一根颈肋、背肋和肩胛骨碎片、一个保存完好但不完整的叉骨、肱骨、第二掌骨、第二指骨第一节、第三指骨第一节、第三指骨第二节、第三手爪骨、第三远侧跗骨、第四远侧跗骨和第二至第五近侧跖骨。正模标本个体很可能在被冲到海里之前就死在了一个古代海滩的岸边。死后,遗骸遭受长时间冲刷,许多骨骼在此期间遗失。[1]
虽然该物种不是水生的,但根据岩层中的菊石判断,尸体所处的环境可能是远洋带。该地区盛产海百合、腹足类、双壳类、腕足类和苔藓虫。[4]沉积发生在浅碳酸盐台地和较深盆地之间的斜坡上。各种各样的划痕、沟槽和条纹表明这只动物的尸体受到了海洋无脊椎动物的啃食。标本代表一个亚成年个体,接近其最大尺寸,年龄估计为24岁。[1]
叙述

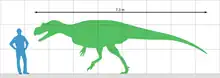
由于遗骸过于零碎,不可能直接测量动物的大小,因此,叙述作者将这些化石与几种体型大致相似的兽脚类进行比较。通过与脆弱异特龙标本MOR 693进行比较,他们保守地得出结论:萨尔特里奥猎龙正模标本个体至少有7到8米长。这将使得该属成为在阿连阶之前已知最大的兽脚类,比侏罗纪晚期的角鼻龙长约25%。与角鼻龙相比,它的体长为730厘米,臀部高度为220厘米,颅骨长度为80厘米。腿骨长度大约是80到87厘米,表明其体重是1160到1524公斤。另一种方法是从已知的前肢长度推断。根据兽脚类动物的肢体比例可推算出后肢长度为198厘米。那么股骨将有822到887毫米长,表明重量为1269到1622公斤。[1]
分类学
萨尔特里奥猎龙的确切分类不能确定,但被视为一种兽脚类。[2][3]达尔·沙索最初将其归入坚尾龙类,[5]但后来认为其可能属于异特龙超科。在这两种情况下,薩爾特里奧獵龍的生存时期都要比其所在演化支的其他成员早大约2000至3000万年。[6]本森在他对大龙属的评论中认为它是腔骨龙超科成员。[7][8]叉骨的存在[6]可能支持将其归入坚尾龙类的分类鉴定,尽管叉骨已被报道来自某种腔骨龙超科。[9][10]
2018年的叙述论文进行了大规模的系统发育分析,发现它是一种基础角鼻龙类,为柏柏尔龙的旁系群。 [1]
| 恐龙总目 Dinosauria |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生态学

萨尔特里奥猎龙化石沉积于一个开放的海洋环境中,尸体可能是从最近的陆地冲入海中,骨骼上存在的Sedilichnus未定种表明无脊椎动物正在清理尸体。[1]萨尔特里奥组的一部分被视为海底斜坡的一部分,该斜坡可能是风暴潮影响下到达的开阔潮下带,且底部洋流恒定。[1]自侏罗纪开始,在赫塘阶至锡内穆阶期间,伦巴第盆地西部存在一片宽阔的陆地区域,在那里形成了温暖潮湿的古气候。[11]萨尔特里奥组发现的恐龙化石可能已经在Arbostora海浪(位于瑞士萨尔特里奥组北部)的作用下被移出该区域。[12]它是一个靠近萨尔特里奥组的出露构造隆起,形成位于Nudo山(东部)和Granoso山(西部)的两个近沉降盆地之间的分界线。[12]它沉积在一个碳酸盐台地上,该台地与从西到东南的其它更广阔区域相连,在北部形成了一个巨大的浅水湾,沉积地层受地垒和构造岩控制。[12]一些所谓的“红土”露头还发现了古土壤,包括萨尔特里奥以西十几公里处的Castello Cabiaglio-Orino。[13][14]这些露头表明,现在的马焦雷湖区域在赫塘阶至锡内穆阶期间曾被森林覆盖,莫尔特拉肖组发现的大型植物碎片证明了这一点。[11]这些植物化石已在Cellina和Arolo(马焦雷湖东侧)之间的区域发现,化石时期与萨尔特里奥组相同。[15]当时的植物群包括本内苏铁目(Ptilophyllum)、南洋杉科(Pagiophyllum)和掌鳞杉科(Brachyphyllum),这些物种皆分布于内陆干热环境中。[15]萨尔特里奥猎龙可能来自附近的陆地,如距发现地较远的Trento地台。如果事实真是如此,那么这种兽脚类很可能是当地最大的掠食者。[1]
参考资料
- Dal Sasso C, Maganuco S, Cau A. 2018. The oldest ceratosaurian (Dinosauria: Theropoda), from the Lower Jurassic of Italy, sheds light on the evolution of the three-fingered hand of birds. PeerJ 6:e5976
- The Theropod Database
- Matthew T. Carrano, Roger B. J. Benson, Scott D. Sampson: The phylogeny of Tetanurae (Dinosauria: Theropoda). (页面存档备份,存于) Journal of Systematic Palaeontology. Bd. 10, Nr. 2, 2012
- Lualdi A. 1999. "New data on the Western part of the M. Nudo Basin (Lower Jurassic, West Lombardy)". Tübingen Geowissenschaftliche Arbeiten, Series A 52: 173–176
- Cristiano Dal Sasso: Dinosauri italiani. Marsilio Editori, Venezia, 2001.
- Cristiano Dal Sasso: Dinosaurs of Italy. In: Comptes Rendus Palevol. Bd. 2, Nr. 1, 2003.
- Roger B. J. Benson: The osteology of Magnosaurus nethercombensis (Dinosauria, Theropoda) from the Bajocian (Middle Jurassic) of the United Kingdom and a re-examination of the oldest records of tetanurans. In: Journal of Systematic Palaeontology. Bd. 8, Nr. 1, 2010, S. 131–146
- Oliver W. M. Rauhut: The interrelationships and evolution of basal theropod dinosaurs (= Special Papers in Palaeontology. Bd. 69). The Palaeontological Association, London 2003
- Larry F. Rinehart, Spencer G. Lucas, Adrian P. Hunt: Furculae in the Late Triassic theropod dinosaur Coelophysis. In: Paläontologische Zeitschrift. Bd. 81, Nr. 2, 2007]
- Ronald S. Tykoski, Catherine A. Forster, Timothy Rowe, Scott D. Sampson, Darlington Munyikwa: A furcula in the coelophysoid theropod Syntarsus. In: Journal of Vertebrate Paleontology. Bd. 22, Nr. 3, 2002
- Jadoul, Flavio, Galli, M. T., Calabrese, Lorenzo, & Gnaccolini, Mario (2005). Stratigraphy of Rhaetian to Lower Sinemurian carbonate platforms in western Lombardy (Southern Alps, Italy): paleogeographic implications. Rivista Italiana di Paleontologia e Stratigrafia, 111(2), 285-303.
- Kalin, O., & DM, T. (1977).(1977): Sedimentation und Paläotektonik in den westlichen Südalpen: Zur triasisch-jurassischen Geschichte des Monte Nudo-Beckens. – Eclogae geol
- Leuzinger, P. (1925). Geologische Beschreibung des Monte Campo dei Fiori u. der Sedimentzone Luganersee-Valcuvia. E. Birkhäuser & Cie.
- Wiedenmayer, F. (1963). Obere Trias bis mittlerer Lias zwischen Saltrio und Tremona:(Lombardische Alpen); d. Wechselbeziehungen zwischen Stratigraphie, Sedimentologie u. syngenet. Tektonik. na.
- Lualdi, A. (1999). New data on the Western part of the M. Nudo Basin (Lower Jurassic, West Lombardy) Tubingen Geowissenschaftliche Arbeiten, Series A, 52, 173-176.