达科他盗龙属
达科他盗龙属(学名:)是种疑似嵌合体的大型驰龙科兽脚类恐龙,[2]生存于晚白垩世的北美,残骸发现于地獄溪組,化石时期约为马斯特里赫特阶,可追溯到中生代末期,让达科他盗龙成为当时仅存的驰龙科之一。化石发现地位于一个含多个物种的尸骨层上,[1]尽管后来发现正模标本和其它归入该属的标本中有部分骨骼属于甲鱼,[3]但还没有对其潜在的非驰龙类亲缘关系作进一步研究;系统发育分析也经常将它放在驰龙科的不同位置[1][4]。
| 达科他盗龙属 | |
|---|---|
 | |
| 骨骼重建 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 獸腳亞目 Theropoda |
| 演化支: | 新獸腳類 Neotheropoda |
| 演化支: | 鳥吻類 Averostra |
| 演化支: | 坚尾龙类 Tetanurae |
| 演化支: | 俄里翁龍類 Orionides |
| 演化支: | 鳥獸腳類 Avetheropoda |
| 演化支: | 虛骨龍類 Coelurosauria |
| 演化支: | 暴盜龍類 Tyrannoraptora |
| 演化支: | 手盜龍形態類 Maniraptoromorpha |
| 演化支: | 手盜龍形類 Maniraptoriformes |
| 演化支: | 手盜龍類 Maniraptora |
| 演化支: | 廓羽盜龍類 Pennaraptora |
| 演化支: | 近鳥類 Paraves |
| 演化支: | 真手盜龍類 Eumaniraptora |
| 下目: | †恐爪龍下目 Deinonychosauria |
| 属: | †达科他盗龙属 Dakotaraptor DePalma et al. 2015[1] |
| 模式種 | |
| †斯氏达科他盗龙 Dakotaraptor steini DePalma et al. 2015[1] | |
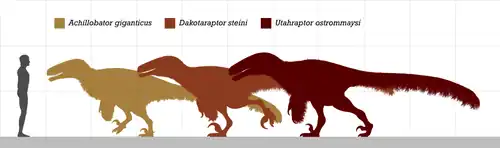
達科他盜龍是已知生存年代最晚的兩種馳龍類之一,另一種是冥河盜龍。達科他盜龍與南方盜龍、阿基里斯龍、猶他盜龍並列為最大型的馳龍科成員。達科他盜龍也是身高最高的馳龍和已知最大的有翅膀的恐龍。
发现与命名
2005年,古生物学家罗伯特·德·帕尔玛在南达科他州哈定县发现了一个骨层,其中含有各种恐龙和非恐龙生物的遗骸,而部分遗骸被认为属于一只大型驰龙科。随后,同一地点又发现了额外的驰龙科遗骸。[5]2015年,模式种斯氏达科他盗龙()由罗伯特·德·帕尔玛(Robert A. DePalma)、大卫·伯纳姆(David A. Burnham)、拉里·迪恩·马丁、彼得·拉尔森和罗伯特·托马斯·巴克等人命名、叙述。属名Dakotaraptor取自化石发现地南达科他州和当地的达科他人以及拉丁语单词raptor(意为“盗贼”);种名steini纪念古生物学家沃尔特·斯坦(Walter W. Stein)。[1]达科他盗龙是2015年开放获取或免费阅读期刊中描述的18个恐龙分类群之一。[6]
正模标本PBMNH.P.10.113.T发现于上地獄溪組的砂层中,可追溯到马斯特里赫特阶晚期,由一具成体的部分骨骼组成,缺少颅骨。它包含一块脊椎、十块尾椎、两块肱骨、两块尺骨、两块桡骨、第一和第二块右掌骨、左手的三只爪子、一根右股骨、两块胫骨、一根左距骨、一根左跟骨、左第二、第三和第四跖骨、右第四跖骨以及右脚的第二和第三爪。一个叉骨后来被排除在标本之外。[7]除了正模标本,在骨层中还发现了属于达科他盗龙的其他骨骼,但形态更为纤细,包括PBMNH.P.10.115.T:右胫骨;PBMNH.P.10.118.T:连接在一起的左距骨和跟骨;以及KUVP 152429:最初被认为是该属的叉骨,但现在也被排除在已知的达科他盗龙化石之外。[7]此外,还有四颗单独的牙齿,标本编号为PBMNH.P.10.119.T、PBMNH.P.10.121.T、PBMNH.P.10.122.T以及PBMNH.P.10.124.T。这些化石是棕榈滩自然历史博物馆收藏品的一部分。其他被提及的化石分别为KUVP 156045:一颗单独的牙齿;[1]以及NCSM 13170:第三根假定的叉骨,后来被确定为不属于达科他盗龙。[7]
叙述
体形

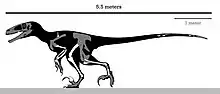
达科他盗龙的体型对驰龙科来说特别大;据估计,成年恐龙的长度为5.5米(18英尺)。[1]2016年,其他估计表明其体长为4.35至6米(14.3至19.7英尺),重220至350千克(485至772磅)。[8][9]
该属的大小接近已知最大的驰龙科——犹他盗龙。然而,达科他盗龙并不具有像后者一样的比例和适应性,而更像恐爪龙这样的小型驰龙科。[1]
鑑定特征
除尺寸较大外,2015年的叙述中还提到了该属的其它显著特征。在第四只脚爪上,用作屈肌腱附属结构的凸起尺寸减小,其外侧朝向尖端的“血槽”在其一半长度上完全封闭,从而形成一个骨的管状结构。脚的第二爪和第三爪下侧有锋利的龙骨。第二只脚爪(即“镰刀爪”)长度相当于股骨的29%。在胫骨上,腓侧嵴(即与腓骨接触的嵴)长而轻巧,高度不超过其9%,上缘末端形如钩状。在第二掌骨与手指接触的两个骨节中,内侧的几乎与外侧的一样大。第二掌骨的外侧只有一条浅槽,以用于连接韧带。当手臂处于平面位置时,在第二掌骨中腕关节和上轴之间的边缘在俯视图中笔直。牙齿后缘每隔5(0.20英寸)有十五至二十个小齿,前缘则有二十至二十七个小齿。[1]
脊柱

背椎高度气腔化,充满海绵骨,显示出许多气室。尾椎中部的前关节突非常细长,估计完整长度为70(28英寸),横跨大约十块椎骨。这使达科他盗龙的尾巴变得格外僵硬。[1]
前肢
像几乎所有其他兽脚类一样,达科他盗龙很可能拥有一个叉骨作为其肩带的一部分。兽脚类动物的叉骨种类繁多,通常与大多数现代鸟类拥有的强烈弯曲的“U”形叉骨不同;最初被认为属于达科他盗龙的叉骨化石呈U形到V形,类似于许多其它驰龙科(如伶盗龙)甚至大型棘龙科兽脚亚目(如似鳄龙)。2015年,维多利亚·阿尔布尔等人的一项研究提出假定的“达科他盗龙叉骨”实际上为甲鱼盔甲的一部分,属于鳖科的一个物种Axestemys splendida的内腹甲。[3]2016年,德·帕尔玛等人认识到,他们在论文中所提及的叉骨实际上不属于达科他盗龙,并将它排除在其种型群之外。[7]
在这篇描述文章中,达科他盗龙的翅膀引起了人们很大的关注。这里,“翅膀”被用作与其功能无关的解剖学描述术语,因为达科他盗龙不会飞。这类似于鸵鸟、鸸鹋和其它不会飞的鸟类的“翅膀”。这意味着手臂上长有长长的羽毛。发现了许多翼骨(肱骨、桡骨、尺骨、三根掌腕骨中的两根以及部分手指),因此翼非常完整。肱骨(即上臂骨)相对较长且纤细,有些向内弯曲。最显著的解剖学特征是在尺骨(前臂骨之一)的下边缘有一排突起,称为尺骨乳状突起或羽茎瘤。在鸟类和其它兽脚类身上,这些突起被作为羽毛附着点。当该结构被发现时,它被认为是这种动物翅膀长时间退化的有力迹象。由于羽茎瘤在化石记录中很少见,古生物学家主要依靠系统发育分析来确定一个物种是否可能有翅膀羽毛——如果进化树中“较高”分支上的一个亲戚有羽毛,而“较低”分支上的一个也有羽毛,那么处于中间位置的一个物种也可能有。达科他盗龙的羽茎瘤表明这种动物具有明显的羽毛,这使它成为具有这种类型羽毛的最大驰龙科。达科他盗龙的羽茎瘤直径约为8—10(0.31—0.39英寸),这表明其附着的羽毛相当大。一根完整尺骨估计可能具有15个这样的羽茎瘤。据测量,尺骨长36(14英寸),另一个下臂骨的半径为32(13英寸)。手骨表明它们的关节几乎不能活动。如果不考虑其原始初级飞羽可能长于其上肢,达科他盗龙的翼展估计约为120(47英寸)。[1]
后肢

总的来说,后肢较为轻盈,有细长的骨头,与犹他盗龙强健结实的后肢相反。与之相比,达科他盗龙更接近某些敏捷、具有灵活性的小型驰龙类,非常适合奔跑和捕食。股骨长度为558(22.0英寸),相对来说比犹他盗龙更短、更轻。相反,胫骨比较长,长度为678(26.7英寸),是已知最长的驰龙科胫骨,比股骨长22%,显示出良好的跑步能力。胫骨肩峰有一个尖角指向前方。它的腓骨嵴终止于一个指向上方的钩状突起,这种情况在整个兽脚亚目中独一无二。距骨和跟骨(脚踝上部的骨头)像斑比盗龙那样融合在一起。跟骨顶部只有一个小型接触面,表明腓骨下端一定很窄。跖骨长度估计为32(13英寸),这使得它相对于后肢的其余部分相当长。[1]
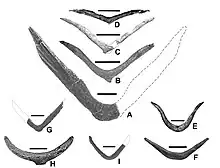
达科他盗龙的脚爪包括一个典型的驰龙科的第二爪,即“镰刀爪”,被用来杀死或压住猎物。它又大又结实,直径为16(6.3英寸);沿外曲线测量的长度为24(9.4英寸)。这相当于股骨长度的29%,相比之下恐爪龙仅为23%。这只爪子横向扁平,并且具有液滴状横截面。屈肌结节是靠近末端的一个大型突起,作为屈肌的附着点——它越大,切割力量就越大。该属的屈肌结节相对于其他已发现的驰龙科来说更大,这可能使它具有该分类群已知成员中最强的砍杀力量。第三只脚爪上的屈肌结节几乎不存在,与其他驰龙类相比,其体积非常小,这表明达科他盗龙很少使用该爪。由于化石仅保存了爪子的骨质核心,它们在生前会被角质层覆盖,延伸出“指甲”,并在末端形成一个尖。第三只爪也向上缩,但是更小,尖端到关节的长度为7(2.8英寸),弯曲长度为9(3.5英寸)。其外侧的凹槽朝向骨隧道的末端,这种情况较为罕见。[1]
分类学
达科他盗龙被归入驰龙科。系统发育分析表明它是驰龙的姐妹群。它们又与犹他盗龙形成了一个分枝,其中阿基里斯龙分支是直系的侧枝。尽管与其它巨型驰龙科具有演化关系,但除恐爪龙、半鸟、阿基里斯龙和犹他盗龙演化支之外,达科他盗龙被认为代表了驰龙类体型增加的第四个例子。[10]
| 真驰龙类 Eudromaeosauria |
| ||||||||||||||||||||||||||||||||||||||||||
莫蒂默·哈特曼(Mortimer Hartman)等人在2019年进行的一项最新研究提出将达科他盗龙归入“半鸟科”。[11]如果这项研究被证明是正确的,那么达科他盗龙可能是已知的最北方的半鸟亚科。
| 半鸟科 Unenlagiidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||

在柯里和埃文斯2019年的系统发育分析中,达科他盗龙再次被发现是一种真驰龙类,尽管作者指出正模标本可能并不来自于同一个体。[12]
| 真驰龙类 Eudromaeosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
在对纳瓦霍战龙的叙述中,史蒂文·贾辛斯基(Steven E. Jasinski)等人(2020年)证实上述观点,但也指出达科他盗龙可能是一具嵌合体,由于该分类单元潜在的嵌合体性质,对这一位置的怀疑过于基础。 [2]
古生物学

第二趾上具有龙骨的巨爪(即“镰刀爪”)被用来杀死猎物,其屈肌结节比犹他盗龙更结实。相反,第三只脚爪的大小相对来说小于其他驰龙类,而且似乎在攻击猎物时没有重要作用。[1]
化石材料中存在两种形态,一种是强壮的,一种是纤细的。一项骨组织学研究表明,这两种形态都属于成年恐龙,因此一些骨骼较轻的构造不是由年龄较小引起的。个体差异或病理学可能解释了这种差异,但最简单的解释是两性异形。目前还不能推断两种形态中哪个是雄性,哪个是雌性。[1]
下臂尺骨上有较宽的尺骨乳状突起或羽茎瘤,是大型羽毛的附着点。成年达科他盗龙因个体太重而无法飞行,但羽毛并没有显示出退化的迹象。其翅膀有多种可能的替代功能,包括保护蛋、展示、恐吓和用镰刀状爪子压住猎物时保持平衡。然而,这些功能并不需要羽茎瘤,叙述作者认为达科他盗龙很可能是一个拥有上述特征的小型会飞祖先的后代。[1]
古生态学

達科他盜龍是地獄溪層中發現的第一種中型食肉動物(除可疑的矮暴龍),長度介于像暴龍这样的巨型暴龍科和像冥河盜龍這样的小型恐爪龍下目之间。[13]其胫骨长于股骨,具有很好的奔跑能力,填补了追捕捕食者的生态位空缺。如果它是一个群居的猎手,成群合作,它可能会捕食更大的食草动物,可与体长6—9(20—30英尺)的未成年暴龙竞争,而成年暴龍是達科他盜龍的天敵。与达科他盗龙共存的恐龙包括很多著名的物种,例如暴龙、三角龙、厚头龙、似鸟龙以及埃德蒙顿龙。[1][15][16]
参考资料
- DePalma, Robert A.; Burnham, David A.; Martin, Larry D.; Larson, Peter L.; Bakker, Robert T. . 2015-10-30 [2022-10-15]. ISSN 1946-0279. doi:10.17161/paleo.1808.18764. (原始内容存档于2015-11-06) (美国英语).
- Jasinski, Steven E.; Sullivan, Robert M.; Dodson, Peter. . Scientific Reports. 2020-12, 10 (1) [2022-10-15]. Bibcode:2020NatSR..10.5105J. ISSN 2045-2322. PMC 7099077
 . PMID 32218481. doi:10.1038/s41598-020-61480-7. (原始内容存档于2022-11-28) (英语).
. PMID 32218481. doi:10.1038/s41598-020-61480-7. (原始内容存档于2022-11-28) (英语). - Arbour, V.M.; Zanno, L.E.; Larson, D.W.; Evans, D.C.; Sues, H. . PeerJ. 2015, 3: e1957. PMC 4756751 . PMID 26893972. doi:10.7717/peerj.1691 (英语).
- Hartman, Scott; Mortimer, Mickey; Wahl, William R.; Lomax, Dean R.; Lippincott, Jessica; Lovelace, David M. . PeerJ. 2019-07-10, 7: e7247. ISSN 2167-8359. PMC 6626525 . PMID 31333906. doi:10.7717/peerj.7247 (英语).
- DePalma, R. A. (Master of Science论文). University of Kansas: 227 pp. 2010 [2022-10-15]. (原始内容存档于2016-08-22) (英语).
- . PLOS Paleo. 2016-01-06 [2020-08-27]. (原始内容存档于2017-07-15) (英语).
- DePalma, Robert A.; Burnham, David A.; Martin, Larry D.; Larson, Peter L.; Bakker, Robert T. . Paleontological Contributions. 2016-12, 2016 (16) [2022-10-15]. ISSN 1946-0279. doi:10.17161/1808.22120. (原始内容存档于2021-11-22).
- Molina-Pérez & Larramendi. . Barcelona, Spain: Larousse. 2016: 275 (西班牙语).
- Paul, Gregory S. . Princeton University Press. 2016: 151 (英语).
- Hartman, Scott; Mortimer, Mickey; Wahl, William R.; Lomax, Dean R.; Lippincott, Jessica; Lovelace, David M. . PeerJ. 2019-07-10, 7 [2022-10-15]. ISSN 2167-8359. PMC 6626525 . PMID 31333906. doi:10.7717/peerj.7247. (原始内容存档于2021-05-23) (英语).
- Currie, Philip J.; Evans, David C. . The Anatomical Record. 2020-04, 303 (4). ISSN 1932-8486. doi:10.1002/ar.24241 (英语).
- Evans, David C.; Larson, Derek W.; Currie, Philip J. . Naturwissenschaften. 2013-11-19, 100 (11): 1041–1049. Bibcode:2013NW....100.1041E. ISSN 0028-1042. PMID 24248432. doi:10.1007/s00114-013-1107-5 (英语).
- Cau, A. . Theropoda. 2015-10-31 [2020-08-27]. (原始内容存档于2016-03-05) (英语).
- Hartman, Joseph Herbert; Johnson, Kirk R.; Nichols, Douglas J. . . Geological Society of America. 2002: 145–167. ISBN 978-0-8137-2361-7 (英语).
- Arens, Nan Crystal; Jahren, A. Hope; Kendrick, David C. . . Geological Society of America. 2014 [2022-10-15]. ISBN 978-0-8137-2503-1. doi:10.1130/2014.2503(05). (原始内容存档于2022-10-15) (英语).