先天免疫系統
先天免疫系统(英語:)又稱為固有免疫、非特异性免疫、非專一性防禦,是生物体非特异性、无记忆性的病原体防御功能,用以抵御病原体入侵的第一道防线,包括一系列的物理屏障系统、固有免疫细胞及固有免疫分子。
先天免疫系统的细胞或分子会非特异地识别并作用于病原体。与後天免疫系統不同,先天免疫系统不会提供持久的保护性免疫,即不产生免疫记忆,且不历经克隆扩增[1],而是作为一种迅速的抗感染作用,存在于所有的动物、植物及真菌中。
炎症反应
炎症反应(发炎)是免疫系统对感染或刺激的第一个回应。它在由受损细胞所释放的化学因子的刺激下产生,并形成一种防止感染扩散的物理屏障。此外,在清除病原后,炎症反应还可以促进损伤组织的愈合。[3]
在炎症反应中产生多种化学因子,包括组织胺、前列腺素、5-羟色胺、白三烯和缓激肽。这些化学因子可以增强痛觉感受器的敏感度、引发血管舒张、召集吞噬细胞和中性粒细胞。随后,中性粒细胞则通过释放细胞因子来召集其他的白细胞和淋巴细胞。
炎症反应会表现出红、肿、发热、疼痛以及可能发生的相关组织器官的功能失常。相应的拉丁文为rubor, tumor, calor, dolor, funtio laesa。
补体系统
补体系统是免疫系统中的一种生化级联反应。它可以帮助或者“补足”抗体本身清除抗原物质或标记抗原物质以待清除的作用。这种级联反应由多种血浆蛋白的相互作用所完成,这些蛋白由肝脏中的肝细胞所合成。这些蛋白完成的工作包括:
补体系统中的各个组成部分具有进化上的保守性,在比哺乳类动物更原始的物种诸如鸟类、鱼类、植物以及部分种类的无脊椎动物都有存在。[4]
先天免疫系统的组成
化学作用
- 汗腺分泌的乳酸、脂肪酸营造了一种酸性环境,抑制细菌的生长。
此外,皮肤在特异性免疫中也起重要作用。 由于皮肤是重要的免疫屏障,因此,烧伤、剥脱性皮炎等原因导致大面积皮肤缺失后,患者将面临严重感染的威胁。
机械作用
生物拮抗作用
细胞

白细胞,又称,是游离于特定的组织器官之外的独立的细胞。它们的功能更类似于独立的单细胞生物,白细胞可以自由地移动并捕捉细胞碎片、外源颗粒或入侵微生物。与许多其他体细胞不同,白细胞没有自行分裂来达到增殖的能力,而是由骨髓中的多能造血干细胞产生。[1]
先天免疫相关的白细胞包括:自然杀伤细胞(NK細胞)、肥大细胞、嗜酸性球、嗜碱性球、涵盖单核-巨噬细胞与嗜中性球吞噬细胞、树突细胞,以及少部分淋巴细胞(γδ T细胞、B1细胞)。这些细胞的作用在于识别和消灭可能导致感染的病原体。[2]
肥大细胞
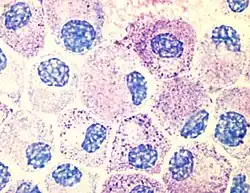
肥大细胞(英語:)实际上是存在于结缔组织和黏膜中的先天免疫细胞,它们与抵御抗原和伤口愈合的作用直接相关,同时也和过敏反应有关。[3]当肥大细胞被激活后,会迅速向细胞外释放其含有的特征性的颗粒(富含组织胺和肝素),以及多种体液调节因子、趋化因子、细胞因子。组织胺会扩张血管,导致炎症的特征性反应,并召集中性粒细胞和巨噬细胞。[3]
吞噬细胞
吞噬细胞(英語:)可以利用胞吞作用吞噬抗原体或其他颗粒。发生作用时,吞噬细胞的细胞膜的局部不断地发生延展和卷曲直到完全裹住胞外的病原体,从而将其转移到胞内。入侵病原体此时被包含在包涵体之中,随后包涵体与溶酶体结合。[2]溶酶体中包含多种的酶和酸性物质,可以杀死并消化病原微生物或颗粒。吞噬细胞通常在体内“巡逻”以搜索抗原,并能够和由其他细胞产生的高度特化的分子信号——细胞因子发生反应。免疫系统中提到的吞噬细胞包括单核-巨噬细胞、中性粒细胞和树突细胞。
在机体正常发育和代谢过程中,这种吞噬作用是很常见的。无论是正常诱导的宿主细胞程序性死亡(也叫做细胞凋亡),抑或由细菌或病毒导致的细胞损伤引起的宿主细胞死亡,吞噬细胞都会负责清除这些死亡的细胞,来保证新的健康细胞的发育和组织的修复。
巨噬细胞
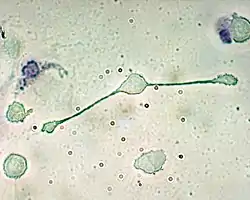
巨噬细胞(英語:)是一种大型白细胞,存在于血管系统中参与循环,也有能力穿过毛细血管的管壁进入到组织间隙中追逐入侵抗原。游走在组织中的巨噬细胞与在血液中的有所不同,称为单核细胞。巨噬细胞作为最有效率的吞噬细胞,可以吞噬相当数量的微生物或其他细胞。[2] 细菌物质分子与巨噬细胞表面的受体的结合会触发巨噬细胞对细菌的包裹作用和杀伤作用。这个过程是通过产生“(呼吸爆发)”的作用,引发活性氧(大多为NO)的释放而发挥作用的。抗原还会刺激巨噬细胞产生趋化因子,使之召唤更多的细胞到感染处来。[2]
嗜中性球
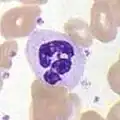
因为在细胞质中存在很多颗粒,嗜中性球(英語:),全称嗜中性粒细胞,与嗜碱性球、嗜酸性球一起并称粒性白细胞,也称作多形核白细胞(polymorphnuclear cells, PMNs)。中性粒细胞的颗粒中含有多种毒素,可以杀死或抑制细菌、真菌的生长。与巨噬细胞类似,中性粒细胞对抗原的攻击也是由“呼吸爆发”所引发的。嗜中性粒细胞是数量最多的一类吞噬细胞,通常占总循环白细胞的50%到60%,并且通常是最先赶到感染区域的淋巴细胞。[3]正常的成人骨髓一天可以产生超过1000亿个中性粒细胞,比急性感染时产量的10倍还多。[3]
树突细胞
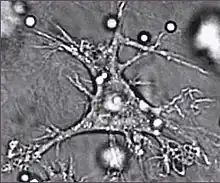
树突细胞(英語:)是存在于组织中并与外界环境相接触的吞噬细胞,通常存在于皮肤(通常称为郎格罕细胞)、鼻粘膜内侧、肺脏、胃脏和肠道之中。[1]树突细胞的命名是因为它很像神经细胞的树突,但二者没有联系。树突细胞是抗原呈現过程中的重要场所,因此也是连接先天和后天免疫系统的纽带.
嗜酸性球和嗜碱性球

嗜碱性球(英語:)激活后,释放的组织胺在清除寄生虫的过程中发挥重要作用,并在过敏反应中发挥作用;[2]激活后的嗜酸性球(英語:)会分泌多种毒素蛋白和自由基,以有效杀灭细菌和寄生虫,但也在过敏反应中造成严重的组织损伤。因此,二者的激活过程十分复杂,以防止不必要的组织损毁。[3]
自然杀伤细胞
自然杀伤细胞(英語:),也简称为NK细胞,主要功能为攻击那些被识别为已感染的宿主细胞,但也同时直接攻击入侵的微生物,例如,NK细胞会攻击并摧毁肿瘤细胞和感染病毒的细胞。这种作用是通过所谓的“自失(missing-self)效应”而发生的,这个效是指被病毒感染的宿主细胞会在细胞表面表现出低水平的MHC I。“天然杀伤”体现在其杀伤那些出现“自失效应”的细胞时无需激活。
γδ T细胞
γδ T细胞是处在先天和后天免疫之间的细胞。一方面,γδ T细胞因为具有产生T细胞受体时的VDJ重排作用而被认为是后天免疫的一部分;而同时γδ T细胞因为具有通过限定TCR受体或NK受体而产生的模式识别受体而被认为是先天免疫的一部分。它可能与非肽抗原的识别有关。
病原特异性
先天免疫系统的不同部分对不同的病原有不同的特异性。
| 病原 | 主要例子[5] | 吞噬作用[5] | 补体系统[5] | 自然杀伤细胞[5] |
|---|---|---|---|---|
| 胞内和胞质病毒 | 有 | 无 | 有 | |
| 胞内细菌 | 有 (特别是中性粒细胞,立克次体无作用) | 无 | 有 (立克次体无作用) | |
| 胞外细菌 | 有 | 有 | 无 | |
| 胞内原生动物 |
|
无 | 无 | 无 |
| 胞外原生动物 |
|
有 | 有 | 无 |
| 胞外真菌 |
|
无 | 有 | 无 |
先天免疫逃避
虽然先天免疫系统的细胞可以有效地阻止细菌在机体内的生长,但是仍有部分病原体成功地进化出了逃避先天免疫反应的机制。[6][7]
绕过先天免疫系统的逃避策略包括细菌在宿主细胞内完成增殖(如沙门氏菌属),以及在表面产生保护性的荚膜来阻止补体和吞噬细胞的裂解作用(如结核杆菌)[8]杆菌属的物种大多是片利共生菌,构成了哺乳动物肠道菌落的大部分。[9]有的物种(如B. fragilis)是条件致病抗原,会导致腹膜腔的感染。这些物种是通过影响吞噬细胞上诱导包被的抗体或者通过模拟宿主细胞来抑制吞噬作用,使得宿主无法将它们识别为外源物质,从而逃避了先天性免疫的。金黄色葡萄球菌可以抑制吞噬细胞对趋化因子的反应能力;而有的物种诸如结核杆菌、化脓葡萄球菌和Bacillus anthracis则可以直接杀灭吞噬细胞来逃避先天性免疫。
细菌和真菌也可以通过形成复合生物薄膜来保护自己;最近的研究表明这种生物薄膜是多种感染的关键所在,比如以囊肿性纤维化为特征的慢性绿脓杆菌感染。[10]
其他形式的先天免疫系统
原核生物的宿主防卫系统
细菌(可能还包括其他的原核生物),利用一种叫做限制修饰系统的防卫机制来抵御来自诸如噬菌体等病原的攻击。在此系统中,细菌通过产生限制酶来攻击并摧毁入侵噬菌体的敌对DNA的特定区域。而自身DNA由于甲基化的作用,得以免除遭到限制酶的攻击。[11]值得注意的是,限制酶和限制修饰系统是仅存在于原核生物中的。
无脊椎动物的宿主防卫机制
无脊椎动物没有淋巴细胞或基于抗体的体液免疫系统。多能的、后天免疫系统很可能是随着脊椎动物而产生的。[12]然而,无脊椎动物的一些免疫机制却可以看做相应的脊椎动物免疫机制的前身。模式识别受体几乎是被所有生物使用的用来识别与微生物抗原相关的分子的一种受体蛋白。Toll样受体是存在于所有体腔动物(包括人)中的一类主要的模式识别受体。[13]补体系统也是存在于大部分生物体内的级联反应系统。只有少数生物如某些昆虫、蟹和蠕虫会使用一种利用prophenoloxidase的补体系统的类似反应系统。[12]
抗微生物肽是先天免疫反应进化过程中相对保守的成分,在所有生物类别都有抗微生物肽。它起到了无脊椎动物的系统免疫功能,如一些昆虫可以产生防御素和天蚕素。
植物的宿主防卫机制
能感染人类的病原生物在分类上处于的纲的很多其他成员都可以感染植物。植物可以通过一系列的复杂代谢反应来形成防御性的化学合成物来抵抗感染或者降低植物对昆虫或食草动物的吸引力。[14]
与无脊椎动物类似,植物既不能产生抗体,也没有T细胞反应,更没有可以通过游走来检测和攻击抗原的细胞。但是,植物却能够做到一些动物罕有的行为来防止感染的扩散,比如隔离或者抛弃那些被感染的部分。[14]
很多植物的免疫反应都包含有遍布植物全身的全身性化学信号。植物通过模式识别受体来检测抗原,并产生基底反应。这个反应产生出的化学信号可以帮助感染隔离作用的产生。当植物的一部分被微生物或者病毒感染后,由特定因子导致的“不兼容反应”使得植物发生局部的超敏反应。并使该处的细胞发生程序性死亡。[15]
“抵抗”蛋白(Resistance proteins, R proteins)是由R基因编码的,广泛存在于植物之中。这种蛋白含有类似于动物先天免疫中模式识别受体和Toll样受体的结构域。植物的系统获得性抵抗(systemic acquired resistance, SAR)通过释放多种化学信使,可以使整个植株获得广谱的抵抗能力。有些化学信使会传遍整个植株,从而使未感染区域产生防卫性的复合物。[14]另外利用RNA干扰机制,植物可以抑制病毒的增殖。[16]
註釋
- Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters. . New York and London: Garland Science. 2002 [2009-03-09]. ISBN 0-8153-3218-1. (原始内容存档于2009-10-03).
- Janeway, Charles; Paul Travers, Mark Walport, and Mark Shlomchik. . New York and London: Garland Science. 2001 [2009-03-09]. ISBN 0-8153-4101-6. (原始内容存档于2009-06-28). .
- Stvrtinová, Viera; Ján Jakubovský and Ivan Hulín. . Computing Centre, Slovak Academy of Sciences: Academic Electronic Press. 1995 [2009-03-09]. (原始内容存档于2007-06-18).
- Janeway CA, Jr.; et al. 6th ed. Garland Science. 2005. ISBN 0-443-07310-4.
- Unless else specified in boxes, then ref is: Lippincott's Illustrated Reviews: Immunology. Paperback: 384 pages. Publisher: Lippincott Williams & Wilkins; (July 1, 2007). Language: English. ISBN 0781795435. ISBN 978-0781795432. Page 172
- Kennedy, Alan. . [2009-03-09]. (原始内容存档于2006-10-10).
- Finlay B, McFadden G. . Cell. 2006, 124 (4): 767–82. PMID 16497587. doi:10.1016/j.cell.2006.01.034.
- Finlay B, Falkow S. (PDF). Microbiol Mol Biol Rev. 1997, 61 (2): 136–69 [2009-03-09]. PMID 9184008. (原始内容 (PDF)存档于2009-03-04).
- Dorland WAN (editor). 30th. W.B. Saunders. 2003. ISBN 0-7216-0146-4.
- Kobayashi H. . Treat Respir Med. 2005, 4 (4): 241–53. PMID 16086598. doi:10.2165/00151829-200504040-00003.
- Restriction Enzymes (页面存档备份,存于) Access Excellence Classic Collection Background Paper.
- Beck, Gregory and Habicht, Gail S. Immunity and the Invertebrates (页面存档备份,存于) Scientific American. November 1996:60-66.
- Imler JL, Hoffmann JA. (2001) Toll receptors in innate immunity. Trends Cell Biol. Jul;11(7):304-11. Review. PMID 11413042
- Schneider, David (2005) Plant immune responses (页面存档备份,存于) Stanford University Department of Microbiology and Immunology.
- Rojo, E.; et al. . Curr Biol. 2004, 14 (21): 1897–1906. PMID 15530390. doi:10.1016/j.cub.2004.09.056.
- Baulcombe D. . Nature. 2004, 431 (7006): 356–63. PMID 15372043. doi:10.1038/nature02874.
參考文獻
- Alberts, Bruce; Alexander Johnson, Julian Lewis, Martin Raff, Keith Roberts, and Peter Walters. . New York and London: Garland Science. 2002 [2018-07-22]. ISBN 0-8153-3218-1. (原始内容存档于2006-08-17).