刃齿虎
刃齒虎屬(Smilodon),舊稱美洲劍齒虎,音譯斯劍虎,其學名在希臘文的意思有「刀一般的牙齒」的意思,大約生存在距今3百萬至1萬年前的北美洲和南美洲。
| 刃齒虎 | |
|---|---|
 | |
| 日本东京国立科学博物馆的致命剑齿虎骨架 | |
絕滅(11130年前) | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 哺乳纲 Mammalia |
| 目: | 食肉目 Carnivora |
| 科: | 猫科 Felidae |
| 演化级: | 假貓級 Pseudaelurus |
| 亚科: | †剑齿虎亚科 Machairodontinae |
| 族: | †劍齒虎族 Smilodontini |
| 属: | †刃齿虎屬 Smilodon Lund,1841 |
| 種 | |
| |
| 異名 | |
|
属异名
种异名
| |
成體約200公斤重,短尾。體型如獅。其犬齒發達,可達17公分。以大型哺乳動物為食;在其獵物滅絕後,斯劍虎也跟著滅絕。
在洛杉磯的拉布雷亞瀝青坑中保存了大量的刃齒虎化石,因此斯劍虎成為最為人所知的史前貓科動物。
分类与种属

刃齒虎属最早是由丹麦自然学家和古生物学家Peter Wilhelm Lund于1841年命名的。他在巴西Minas Gerais州的Lagoa Santa镇附近发现了现在称之为毁灭刃齒虎的动物的化石。
有一些此属的种被描述过,但一般只三个种得到承认。[1]
- 纤细刃齒虎(Smilodon gracilis),是体型最小(体重在55到100公斤)[2]和发现最早的种,生活在距今大约250万年到50万年间。它可能是由北美的巨颏虎(Megantereon)进化而来,并成为其他种剑齿虎的祖先。
- 致命刃齒虎(Smilodon fatalis),生活在距今160万年到1万年间。在北美取代了纤细刃齒虎的地位并在南北美洲生物大遷徙中进入了南美西部。它的体型介于纤细剑齿虎和毁灭剑齿虎(Smilodon populator)之间,大约与现存的最大的猫科动物西伯利亚虎相当[2]。它们肩高可达1米,体重在160到280公斤之间。人们有时会提到加州刃齒虎(Smilodon californicus)和Smilodon floridanus,但通常它们被认为是致命剑齿虎下的亚种。
- 毁灭刃齒虎(Smilodon populator,“Smilodon the Devastator”),生活在距今100万年到1万年间,在南美东部出现,是剑齿虎亚科下体型最大的种。它比它的近亲致命剑齿虎和纤细剑齿虎庞大得多,拥有宽大的胸部和前肢,是已知的剑齿猫科动物中最大的。它肩高大于1.4米,平均身长2.6米,尾长30厘米,体重大约在360到470公斤之间,是已知最重的猫科动物之一。[2][3]
毁灭刃齒虎的上犬齿可达30厘米并伸出上颚达17厘米之长。遗传学证据表明毁灭刃齒虎和该属的其他成员在约1400万年到1800万年间与现存猫亚科动物的祖先已经在进化上分离。[4]
毁灭刃齒虎也是从位于Ultima Esperanza的著名洞穴地发现并为人所知的,一同发现的还有保存完好的与之有关的内源DNA。[4]
系统发生学
| 貓科 Felidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
解剖
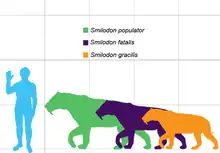
刃齒虎是地球上生存过的大型猫科动物之一。身型庞大的毁灭刃齒虎可能拥有超过500公斤的体重。[2]
一只发育完全的刃齒虎依种类不同体重在55公斤到470公斤之间。它的尾巴很短、四肢有力、颈部健壮、犬齿极长。剑齿虎比现今的所有猫类都要强壮,大概跟熊差不多。它的背侧腰部按比例缩短,与现代豹属的其他动物相比,其后肢比前肢要短,说明刃齒虎在奔跑速度上并不在行。
本属体型最大的种当为南美的毁灭刃齒虎(Smilodon populator)。它的肩部高于臀部,腰部向后下方延伸,乍看之下很像鬣狗;而致命刃齒虎(Smilodon fatalis),平坦的后腰让它们看起来更像现今的猫科动物。但要注意的是,虽然毁灭剑齿虎的前肢较长,但整体的构造十分强壮。和现今的猫科动物和致命剑齿虎相比,它的前臂要比上臂短。这表明它的前肢适于用力而非奔跑,且在捕食时会发挥出巨大力量。
四肢

刃齒虎的四肢比其他猫科动物要短且强壮,其前肢发育良好的屈肌和伸肌让它能轻易扑倒大型猎物。它的后肢拥有强大的内收肌,能帮助刃齒虎在与猎物的搏斗中保持稳定姿态。与大多数猫科动物一样,它的爪子也是可伸缩的。[5]
牙齿和下颌

剑齿虎最负盛名的就是它那长长的犬齿,是剑齿猫科动物中最长的。毁灭剑齿虎(Smilodon populator)的犬齿最长可达28公分。
尽管比其他的大型猫类都要强壮,刃齒虎的咬合力并不强。现今的大型猫类拥有很大的颧弓,而相比之下刃齒虎的颧弓要小。这限制了颞肌的厚度和力量,进而限制了剑齿虎的咬合力。对剑齿虎狭窄下颌的分析表明它所能产生的咬合力量仅相当于一头狮子的三分之一。[6]似乎有规律表明越长的犬齿预示着越弱的咬合力。然而对犬齿的抗弯曲能力(犬齿被外力弯曲但不致折断)和割咬能力的测试表明,与现今的大型猫类相比,在产生同样割咬能力的条件下,剑齿虎的犬齿能承受更大的弯曲力。[7]另外,分析表明剑齿虎能张开下颌到120度,狮子只能到60度。[8][9]
有人认为刃齒虎的较为薄弱的颞肌(控制咬合力的肌肉)并不是设计用来杀死猎物。它们杀死猎物的方式是先用强大的颈部和前肢肌肉制服猎物,接着张开大嘴包围住猎物的咽喉,插入锋利的犬齿割断血管气管致其死亡。根据这个假说,刃齒虎杀死猎物是依靠颈部的伸肌而非下颌的肌肉。
食物及捕猎
刃齒虎很可能捕食多种大型动物,包括美洲野牛、貘、鹿、美洲驼、擬駝、马以及大地獭等。如同似剑虎一样,剑齿虎也有可能捕食未成年的乳齿象和猛犸象。它们甚至可能会攻击史前人类,尽管这一点没有实证。在拉布雷亚沥青坑中有数以百计的剑齿虎被困于沥青之中,很可能是因为它们当时正在捕食同样被困住了的猛犸象。洛杉矶县自然历史博物馆保存有很多剑齿虎的完整骨架化石。

现代的大型猫科动物主要通过切断猎物的气管来杀死猎物,这一过程通常需要数分钟。剑齿虎下颚相对薄弱的肌肉可能不能派作此用,同时它长且脆的犬齿及单薄的颅骨可能会在与激烈反抗的猎物的争斗中受伤。一项2007年的研究表明剑齿虎很可能是通过强大的上肢先将猎物牢牢按倒在地,然后用犬齿切割猎物咽喉,进而割断颈动脉及气管以迅速杀死猎物。[10]这项研究的作者声称这种猎杀方式能让剑齿虎相对于现代猫科动物更适合于对付大型猎物,但同时也让剑齿虎更依赖于大型猎物。这种高度特化的捕猎方式可能加速了剑齿虎的灭绝。剑齿虎相对笨拙的身形和过长的犬齿让它们难以捕到体型小并移动迅速的猎物,这样一旦其生存的生态系统因故发生变化造成大型动物稀少,则剑齿虎将会面临巨大困境。[6]后续的一项研究也支持这种观点,[5]这项研究发现剑齿虎相对现代大型猫科动物,其上肢更适于牢牢控制猎物。
曾有这样的研究:在非洲,研究者们播放受困动物的叫声,借此观察食肉动物对此的反应。这种研究方法也被应用到了对拉布雷亚沥青坑中物种的分析上。利用这种方法,人们发现受困的动物——比如被困在沥青坑之中的动物——发出的叫声往往会吸引群体性的捕食者如狮子和斑鬣狗,而非单独行动的捕食者。鉴于在拉布雷亚沥青坑中发现的捕食者主要是剑齿虎和群居性的恐狼,这提示剑齿虎可能和恐狼一样,也是一种群居性动物。[11]有学者据此提出,如果真是如此,那么剑齿虎夸张的长犬齿可能更多的会用来进行群体成员之间的信号传递或交配。[12]然而,剑齿虎的犬齿并无两性异形,这一点不支持犬齿是用来传递信号或交配的观点。
群居假说
一直以来短剑虎被认为是独自生活的,虽然这个设想并无事实依据,但在长达150年的时间里人们相信这是真的。但当与之不符的证据被发现,古生物学家们开始重新审视这个古老的观点。
在推断灭绝已久的动物的生活行为时,很多结论是通过参考现存的且与灭绝动物处于大致相等生态位的动物得来的。第一个关于群居短剑虎的假说是通过加州剑齿虎的骨层研究提出的,并且这个假说几乎全部来自于对现存物种的研究。
群居假说的一个证据来自于对拉布雷亚沥青坑中动物数量、生态和食肉动物行为的比较,这项比较提出不光剑齿虎,可能其他的短剑虎都是集体捕猎的群居性动物。
来自拉布雷亚沥青坑的证据

位于洛杉矶的拉布雷亚沥青坑是世界上剑齿虎化石最集中的地点之一。此处原本是一个湖,湖水之下是厚厚的沥青,再往下就是化石层。古代的动物来到湖边喝水时,像现代的动物一样有时会把身体浸到水里清洁皮肤或是摆脱寄生虫,这时它们就陷入了湖底的沥青中,而每一次挣扎都让它们陷得更深。动物们并不会立即死亡,它们往往在好几天之后才死于饥饿或休克。它们挣扎和呼救会吸引猎食者来到湖边并同样被困住。这样拉布雷亚沥青坑就成为了一个猎食者的陷阱。一头被困的美洲野牛往往会吸引数倍的捕食者。据统计坑中捕食者与猎物的数量比例大约为九比一。

剑齿虎总共约有2000个被复原的个体,提供了约13000标本,在拉布雷亚沥青坑中的化石丰富程度仅次于恐狼(约20000标本,4000复原个体)。[13]剑齿虎总被认为是独居动物,对它的描述就如秃鹫一般:众多单独的动物争夺一具猎物尸体,彼此以獠牙示威。这种想法预示捕食者和猎物的数量比会相当高。
生态和数量
《The Dinosaur Heresies》一书的作者,Robert Bakker,是首先运用生态学中营养级的概念来支持某一假说的学者之一。[14]他声称恐龙是热血动物并解释说“保持体温恒定是十分耗能的,一头一百磅的看门狗(加上小狗)一年需要一千磅的狗粮。但如果不保持恒定体温则耗能要少许多。一只一百磅的蜥蜴(加上孵化的小蜥蜴)一年只需一百磅的粮食就足够了。”[15]所以,理论上一千磅的食物能支持数量十倍于热血动物的冷血动物。通过确定恐龙猎食者和猎物的比例,人们发现这个比例符合热血动物的推定。这种比例也说明为什么一个生态系统中捕食者的数量总是会比猎物少。对一个生态系统的准确研究表明,猎食者和猎物九比一的比例是不可能的。[16]这个数字得有个解释。

声音当然是一个因素。当一只动物被困在沥青坑里,它在恐慌和垂死之际必然会发出哀嚎。这哀嚎或是向同类呼救,或仅是无意义的挣扎。但这哀嚎也会吸引远处的捕食者,提醒它们这里有一顿唾手可得的大餐。
这个解释隐含着一个奇怪的推论:被哀嚎吸引而来的动物是群居性的。[17]在东非,非洲野犬的叫声通常意味着它们由于杀戮而感到兴奋。狮子则并不是人们想象的那样是勇敢的捕食者,它们是机会主义者。狮子从鬣狗那偷来的猎物和它们自己捕猎得到的相比只多不少。狮子如果听到可能意味着食物的叫声,它们是会前往一探究竟的。独自行动的豹则从来不会去理会这些声音。其他的一些独居的猫科动物,如薮猫和猎豹,也不会对这些声音感兴趣。东非的另一种半群居的、只在哺乳时结伴而行的犬科动物,豺,只在食物非常缺乏时才会去冒险。
食肉动物对声音的反应
一项针对群居捕食者对声音反应的研究指出拉布雷亚沥青坑中大量剑齿虎的出现是因为它们群居。[18]研究者播放垂死的食草动物、狮子和鬣狗的叫声并记录下循声而来的动物。这些食肉动物被分为四类:大的(体重21公斤以上)群居性动物、大的独居动物、小的(21公斤以下)群居性动物和小的独居动物。研究者比较了对声音感兴趣的动物的年龄和沥青坑中剑齿虎的年龄(从掉落的牙齿估计而来)。他们在南非的Kruger国家公园、坦桑尼亚的Serengeti国家公园及周围保护区的实验结果如下表A和B列所示。
| 类别 | A 非洲生态系统中个体数量 |
B 对所播放声音感兴趣的个体数量 |
C 若剑齿虎独居, 北美沥青坑中个体数量 |
D 若剑齿虎群居, 北美沥青坑中个体数量 |
|---|---|---|---|---|
|
大体型且群居 |
2 % |
84 % |
53 % |
87 % |
|
大体型且独居 |
1 % |
1 % |
36 % |
2 % |
|
小体型且群居 |
8 % |
15 % |
8 % |
8 % |
|
小体型且独居 |
89 % |
0 % |
3 % |
3 % |
困境中猎物的哀嚎能吸引附近区域中捕食者的注意。如果每一个捕食者都循声而来,那么可以想见会有激烈的竞争。体型小且群居的豺,体型大且独居的豹在这样的竞争中可能会吃亏,但它们还是可能会小心翼翼地在附近查看一番。豺就是如此,若是有大的捕食者正在进食,它们会在周围盘桓,小心推进,希望在对方鼻子底下偷一杯羹或是等对方饱食而去后再去吃残羹冷炙。体型小且独居的动物,如獾、薮猫、麝猫、非洲麝猫等,它们在这样的竞争中不仅仅可能被杀,更可能成为别人食物的一部分。所以它们对这种哀嚎的反应不是置之不理,而是当作危险信号,它们会及时离开以躲避大群即将到来的大型捕食者,这也是表格所显示的。大体型且群居的狮子,得益于它们的力量和数目,在竞争中最为有利。其他能与狮子分庭抗礼的就是斑鬣狗了。斑鬣狗不仅在常规猎杀中常常挑战狮子,它们也会被动物的哀嚎所吸引。其他的捕食者往往会避开斑鬣狗和狮子的争斗以避免自身受到牵连。
为了进一步说明情况,拉布雷亚沥青坑中的遗骸被按照与非洲生态系统实验中相同的方式分类。沥青坑中的样本与非洲生态系统中的种群并不具可比性,但还是能用类似的方法去分析。如果剑齿虎是独居动物,那么结果就是上表中的C列,如果剑齿虎是群居动物,那么结果就是上表中的D列。这两列的结果,和B列中的结果相比,很明显地支持剑齿虎的群居假说。
群居的好处之一是能在和其他群居捕食者,如美洲狮和恐狼的竞争中占得优势。很多群居社会性动物都存在雌雄异态,但剑齿虎不是这样。雌性和雄性剑齿虎的犬齿长度相仿,这提示剑齿虎的群居社会结构可能更类似于现代的狼而非狮子。这也提示剑齿虎可能和豺一样是一夫一妻制,而不像狮子的一夫多妻制。
受伤的骨骼和古生物病理学
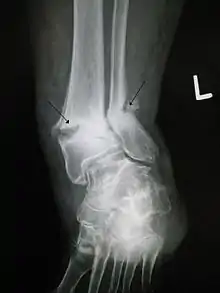
来自化石本身的证据也支持群居假说。当一只动物的活动超过身体承受范围时,受伤是不可避免的。剑齿虎的骨骼经常显示骨折和变形,脱离骨骼的肌肉也比较常见。猎杀大型猎物可能会导致这样的损伤。令人感兴趣的是这些损伤愈合了。从骨骼上脱落的肌肉愈合,再脱落,再愈合,再脱落。这种循环让剑齿虎的骨骼上由于钙的大量沉积而长出肿块,骨折虽然恢复得很差,但毕竟是恢复了。[19]独居动物,如猎豹,一个小小的损伤就可能让它无法捕猎直到饿死。身体健全的情况下捕猎已经是很困难的了,身体如果在饿死之前还没有修复损伤,那么骨折便永远不会愈合了。剑齿虎看来承受了巨大的伤害,但它挺过了修复的阶段并恢复了捕食。剑齿虎亚科下最大的短剑虎属,同剑齿虎一样,常常有磨损的半截犬齿,表示在犬齿断掉之后还常常使用。[20]
化石骨骼的变形里的一种特殊情形是在一只接近成年的剑齿虎化石上发现的骨盆的变形,[21]和所显示的外伤性骨化性肌炎。这些损伤让这只动物完全无法行动直至愈合完全,但即使愈合之后它也很可能只能一瘸一拐地走路。所显示的肌肉损伤非常严重,很可能会造成皮肤下的大量淤血。在这几个月的恢复期间,这只动物会非常疲惫和痛苦。它需要别的群体成员给它带来现成的食物,这些食物很可能来自它修养之处附近的猎场。在母狮中曾观察到过长达九个月的这种行为(Schaller, 1972)。
一项针对骨骼的研究试图使用古生物病理学的方法,根据5000个变形的化石样本来推测这些动物的生活。[22]古生物病理学是根据化石记录上由于断裂、张力、剥落、肿胀、钙沉积、咬痕等造成的形变来研究伤病古生物的学科。这些化石上的形变显示了很多能让剑齿虎衰弱的损伤,如断裂和磨损的犬齿、剥落的门牙和磨牙、关节炎、由真菌感染引起的溪谷热、严重感染的臀部、骨折的脖子等。[23]反复受到损伤的肌肉会在恢复时在骨骼上留下节瘤,通过观察哪里的损伤最多,可以推断出哪些肌肉用的最多,进而了解剑齿虎如何捕猎。
灭绝
剑齿虎在更新世晚期,大约公元前10000年时灭绝,这个时期还有很多其他更新世巨型動物群的大型食草和食肉动物灭绝。
这时期到达北美的史前人类猎杀了很多大型动物,被认为导致了这次大灭绝。也有观点认为是冰河期的结束导致了这次灭绝,因为冰层的消失会使植被发生变化。草原延伸了,夏季更热更长了,北美部分地区陷入干旱。但这种观点无法解释为什么剑齿虎和其他大型动物在之前的很多次冰河期间隔中能生存下来,却在这次冰河期末期在北美和南美突然灭绝。
参考文献
- A. Turner: The big cats and their fossil relatives. Columbia University Press, 1997. ISBN 978-0-231-10229-2
- Christiansen, P.; Harris, J. M. . Journal of Morphology. 2005-12, 266 (3): 369–384 [2011-08-02]. PMID 16235255. doi:10.1002/jmor.10384.
- Sorkin, B. . Lethaia. 2008-04-10, 41 (4): 333–347 [2011-08-02]. doi:10.1111/j.1502-3931.2007.00091.x. (原始内容存档于2020-03-29).
- Barnett, Ross; Ian Barnes, Matthew J. Phillips1, Larry D. Martin, C. Richard Harington, Jennifer A. Leonard, and Alan Cooper. . Current Biology. 9 August 2005, 15 (15): R589–R590 [2007-06-04]. PMID 16085477. doi:10.1016/j.cub.2005.07.052. (原始内容存档于2017-11-02).
- Meachen-Samuels JA, Van Valkenburgh B. (2010) "Radiographs reveal exceptional forelimb strength in the sabertooth cat, Smilodon fatalis." PLoS One 2;5(7):e11412.
- Jeff Hecht. . New Scientist. 1 October 2007 [2012-03-27]. (原始内容存档于2015-05-23). 这项研究使用了一种称作有限元分析的在工程领域广泛使用的技术。
- Christiansen, P. . Zoological Journal of the Linnean Society. October 2007, 151 (2): 423–437 [2012-03-27]. doi:10.1111/j.1096-3642.2007.00321.x. (原始内容存档于2012-10-21).
- For the lion's gape, see Martin LD. . Janis CM, Scott KM, Jacobs LL (编). Vol. 1. Cambridge University Press, 1998-2008: 234. ISBN 0-521-25519-2 请检查
|isbn=值 (帮助). - . [2012-03-27]. (原始内容存档于2021-02-05).
- McHenry, C.R., Wroe S., Clausen, P.D., Moreno, K. and Cunningham, E. . PNAS. October 2007, 104 (41): 16010–16015 [2012-04-08]. PMC 2042153
 . PMID 17911253. doi:10.1073/pnas.0706086104. (原始内容存档于2020-03-29).
. PMID 17911253. doi:10.1073/pnas.0706086104. (原始内容存档于2020-03-29). - Carbone C, Maddox T, Funston PJ, Mills MG, Grether GF, Van Valkenburgh B. . Biol. Lett. February 2009, 5 (1): 81–5. PMC 2657756 . PMID 18957359. doi:10.1098/rsbl.2008.0526.
- . [2012-04-08]. (原始内容存档于2012-04-28).
- . [2012-04-21]. (原始内容存档于2019-04-25).
- . [2012-04-23]. (原始内容存档于2020-12-05).
- Bakker, Robert. . Kensington Publishing Corp. 1986. ISBN 978-0-82175-608-9.
- . [2012-04-23]. (原始内容存档于2012-05-11).
- . (原始内容存档于2012-03-24). Authors list列表中的
|first1=缺少|last1=(帮助) - . [2012-06-06]. (原始内容存档于2016-04-12).
- . (原始内容存档于2008-03-17).
- Switek, Brian. . [2012-06-10]. (原始内容存档于2012-11-07).
- (PDF). (原始内容 (PDF)存档于2012-03-28).
- Mydans, Seth. . The New York Times. 1989-09-26 [2012-06-10]. (原始内容存档于2019-06-19).
- Biederman, Patricia. . Los Angeles Times. 1989-06-11 [2012-06-10]. (原始内容存档于2016-03-08).
 Smilodon californicus
Smilodon californicus
國立自然史博物館, 華盛頓特區 现存日本上野森博物馆的致命剑齿虎(Smilodon fatalis)头骨
现存日本上野森博物馆的致命剑齿虎(Smilodon fatalis)头骨 毁灭剑齿虎(Smilodon populator)复原图
毁灭剑齿虎(Smilodon populator)复原图