蜜蜂屬
蜜蜂屬(學名:Apis)是膜翅目、蜜蜂科下的一類真社會性昆虫,会採食花粉和花蜜並釀造和储存蜂蜜,利用蜂蜡来建造长期居住的群体巢穴。蜜蜂屬内已被认可的物种仅有8个,其中最知名的是西方蜜蜂(Apis mellifera),在全球被广泛饲养,用以生产蜂蜜制品,飼養蜜蜂者稱為蜂農。
| 蜜蜂屬 化石时期: | |
|---|---|
 | |
| 正在吸食花蜜的西方蜜蜂。牠的體表密被細毛,会产生轻微的静电,使花朵雄蕊中的花粉吸附在牠的身上,並通过後續的活動將花粉傳送至其他同種花朵的雌蕊中。 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 节肢动物门 Arthropoda |
| 纲: | 昆虫纲 Insecta |
| 目: | 膜翅目 Hymenoptera |
| 科: | 蜜蜂科 Apidae |
| 演化支: | Corbiculata |
| 族: | 蜜蜂族 Apini Latreille, 1802 |
| 属: | 蜜蜂屬 Apis Linnaeus, 1758 |
| 模式種 | |
| 西方蜜蜂 A. mellifera Linnaeus, 1758 | |
| 亞屬和種 | |
| |
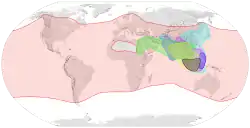 | |
| 各种蜜蜂的分布范围: 西方蜜蜂 東方蜜蜂 小蜜蜂 | |
蜜蜂只是2万个已知蜂種當中的一小部分[2]。其他部分种类的蜂(包括无蛰蜂)也生产和储存蜂蜜,但只有蜜蜂属的成员才被视作真正的蜜蜂。对于蜂的研究,包括对蜜蜂的研究,被称为蜂學。
蜜蜂是農糧作物的重要授粉者,不僅是生態系不可或缺的角色,更維繫全球糧食安全。2018年起,聯合國將每年5月20日訂為世界蜜蜂日,以提高大眾對蜜蜂及相關議題的重視。[3]
词源与名称
蜜蜂属的学名 Apis 是拉丁语,意为“蜂”[4]。现代英语词典里面可能会将Apis解释为“honey bee”或者合成词“honeybee”。昆虫学家罗伯特·斯诺德格拉斯(Robert Snodgrass)声称正确的用法应该是前者,因为“蜜蜂”是“蜂”的一个种类或类别,把两个单词合成一个是不正确的,反例是“蜻蜓(dragonfly)”与“蝴蝶(butterfly)”,后两者并不是苍蝇(fly)[5]。(英文缩写:ITIS)以及生命之树(英文缩写:ToL)中使用的名称为“honey bee”[6][7][8]。然而,合成词用法(honeybee)在自然语言的正词法规则中逐渐地固定下来,这种固化并不见得遵守某种规则。
日常所说蜂王和蜂后是同一个意思,均为蜂后。
起源、分类学与分布
蜜蜂的起源中心似乎位於南亚与东南亚(包括菲律宾)地区,因为除西方蜜蜂以外的现生蜜蜂均原产於该地区。值得注意的是,一些最初谱系的现存典型代表(小蜜蜂和黑小蜜蜂)在该地区都有牠们的起源中心[9]。
蜜蜂屬最早的化石產自欧洲大陆始新世和漸新世之交(約3400万年前)的岩層中。这些化石并不能充分表明欧洲即是蜜蜂属的起源地,只能说明当时的蜜蜂已经活跃于欧洲。作为蜜蜂的疑似起源地,南亚地区发现的相關化石很少,得到深入研究的更是寥寥無幾。
自人类时代以来,新大陆上没有发现蜜蜂存在,直到欧洲人引进了西方蜜蜂。新大陆上只有一个化石被记录在案——古北美蜜蜂 (Apis nearctica),来自内华达州的单个1400万年前的标本[10]。
现代蜜蜂的近亲——比如熊蜂(學名:Bombini;英文:bumblebee)和无蛰蜂(學名:Meliponini;英文:Stingless bee)——在一定程度上也是社会性昆虫,而且社会性这一支序分類學特征似乎在蜜蜂属的形成之前就出现了。在蜜蜂属的现存成员中,偏基底的种类会構築单个裸露的蜂巢,而較晚出現的种类则会在洞穴中構築多个蜂巢,这也大大促进了对牠们的驯化。
历史上大多数蜜蜂种类都被人类养殖过,或者至少在其原生环境周围被土著人开采蜂蜜与蜂蜡。只有两种蜜蜂真正意义上地被驯化——西方蜜蜂和印度蜜蜂(Apis cerana indica)。西方蜜蜂的养殖能追溯到至少埃及金字塔建造的时代,也只有这个蜂种被广泛地培育到了其原生环境范围以外。
蜜蜂屬為蜜蜂族(Apini)中唯一现存的属,并分为三个亞屬:小蜜蜂亚属(Micrapis)、大蜜蜂亚属(Megapis)與蜜蜂亚属(Apis,含家养蜜蜂及其近亲)[11][12]。
小蜜蜂亚属

小蜜蜂和黑小蜜蜂(Apis andreniformis)是南亚与东南亚的小型蜜蜂。牠们的巢穴非常小,裸露在树上或灌木中。牠们的蜇针通常无法穿透人体皮肤,因此人们可以在最小的防护措施下接近其蜂巢与蜂群。牠们大部分并存与同一地区,但不会杂交而成混合种。尽管牠们演化得非常不同,而且可能是异域成种,然而牠们的分布后来发生了融合。
小蜜蜂分布得更广,黑小蜜蜂更具侵略性,通常只有前者会酿造蜂蜜。牠们是蜜蜂之中最古老的现存谱系,可能是在巴尔顿期(4千万年前或者稍晚些)从别的谱系分离而出,但是两者在新近纪之前的很长一段时间内似乎都还没有互相物种分离。[12]小蜜蜂较之其姐妹物种具有更小的翼展。[13]小蜜蜂全身黄色,除了其工蜂的小盾片部位是黑色。[13]
大蜜蜂亚属
大蜜蜂亚属中有兩个种。牠们通常会筑单个或少数几个巢穴,巢穴裸露于高高的树枝、悬崖上面,有时候也会在高层建筑上。牠们可以变得非常凶猛。人类的“蜂蜜猎人”会定期地掠夺牠们的蜂蜜,蜂群一旦被激怒可以轻而易举地将一个人类蛰至死亡。
大蜜蜂亚属包含以下种以及亚种:
- 大蜜蜂(Apis dorsata)原产于并且广泛分布于南亚于东南亚的大部分地区。
- 印度尼西亚大蜜蜂被分类为大蜜蜂的印度尼西亚亚种,或者被分类为独立的种,在后者情况下小舌亚种(A. d. breviligula)和其他谱系可能也必须被认为是独立的种了。[14]
- 喜马拉雅大蜜蜂(Apis laboriosa)最初是大蜜蜂的亞種,1980年依形態上的差異分成独立的种,后来1999年被恩格爾(Michael S. Engel)重新歸為是大蜜蜂的亚种[11](基于生物学物种概念),尽管那些采用遗传学物种概念的作者们建议牠应当被归为一个独立的种,2020年根據特定站點研究觀察顯示,本種確實一直和大蜜蜂共存著,在外形上跟大蜜蜂所差無幾,但沒有和大蜜蜂有雜交的情況發生,因此再度成為獨立的種,也證實了本種的和大蜜蜂有生殖隔離的情形。[12]牠们只生活在喜马拉雅山脉,却具有广泛的行为适应性,使其可以在高纬度地区的低温外部环境中筑巢。牠们是现今体型最大的蜜蜂。
蜜蜂亚属

蜜蜂亚属包括三个或者四个种,分别是沙巴蜂(Apis koschevnikovi)、東方蜜蜂和西方蜜蜂。其中,来自婆罗洲的沙巴蜂是一个明确的独立物种,牠们可能是由穴居蜜蜂在岛屿上的初次定居而衍生出来的。养殖蜂种是東方蜜蜂和西方蜜蜂。西方蜜蜂的遗传学机制尚未明晰。
| 东方蜜蜂 | 西方蜜蜂 | |
|---|---|---|
| 工蜂 | 上唇基具三角斑,后翅中脉分叉,个体较小 | 上唇基无三角斑,后翅中脉无分叉、个体较大 |
| 头向巢门扇风,飞翔速度快,能在7℃气温中采集,失王后工蜂较快产卵,产雄性卵 | 头向巢门外扇门,多数西方蜜蜂飞翔速度不及东方蜜蜂。低温采集能力不及东方蜜蜂。失王后多数西方蜜蜂工蜂产卵慢,其中海角蜂工蜂能产出雌性卵 | |
| 蜂王 | 生殖道口无瓣膜突,一侧卵巢管为100–120 条 | 生殖道口有辦膜突,一侧卵巢管150–200条 |
| 每日产卵量为400–1000粒, 产卵盛期1–1.5年 | 每日产卵量为800-1500粒,产卵盛期2–3年 |
东方蜜蜂

東方蜜蜂(Apis cerana)是南亚与东亚的传统養殖蜜蜂。牠们被驯化为印度蜜蜂(A. c. indica)、中國蜜蜂(A. c. cerana)、黑中國蜜蜂(A. c. heimifeng)和日本蜜蜂(A. c. japonica)等亞種,养殖在蜂房中,与西方蜜蜂类似,但是规模更小。东方蜜蜂与東南亞分布的亞種,如婆罗洲蜜蜂(A. c. nuluensis)之间的关系尚未得到满意的解答;最新的假说认为,牠们确实是不同的物种,但是东方蜜蜂是一个並系群,可能包含了一些考慮独立出來成种的亞種。[12]
西方蜜蜂
西方蜜蜂(Apis mellifera),最常见的家养蜜蜂[16],是昆虫中第三个完成基因组测绘的。牠们似乎起源于非洲西部的热带,并从那里扩散到欧洲南部以及往东进入亚洲直达天山山脉。在世界的不同地方,牠们有不同的名字——“欧洲蜜蜂”“西方蜜蜂”或者更常用的“蜜蜂”。许多亚种已经适应了当地的地理与气候环境,此外,一些混种株系——比如巴克法斯特蜂——孕育了出来。亚种之间或甚至株之间的行为、颜色及解剖学特性可以千差万别。
西方蜜蜂和东方蜜蜂在中新世晚期分化,可能是由於中东地區的沙漠化導致二者的共同祖先東西分隔,基因流動就此中斷,最終形成向东非发展的西方支系和向热带亚洲发展的东方支系。
西方蜜蜂的亚种的多样性可能是更新世早期大规模輻射適應的结果,上一个冰河时期的气候与栖息地的变化对此推波助澜。人类几千年来对西方蜜蜂的集中饲养——包括杂交与外部引进——显然加速了牠们演化的速度,并将其 DNA 序列充分混淆,以至于这许多西方蜜蜂的亚种之间究竟有何关联也已经说不清楚了。[12]
西方蜜蜂并非原产于美洲,因此欧洲探险者与殖民者到达美洲时牠们并不存在。然而,其他当地蜂种却被土著人养殖并交易着。1622年,欧洲殖民者将欧洲黑蜂带到了美洲,随后带来了意大利蜂及其他种类。殖民时代以来,许多依赖于蜜蜂授粉的作物相继被引进。逃逸的蜂群(被称为野蜂)迅速扩散,占领北美大平原的速度通常比殖民者还要快。自然条件下蜜蜂并没有穿越落基山脉,但牠们在19世纪40年代末期被摩门教徒的先驱们运到了犹他州,并在19世纪50年代经船运到达加利福尼亚州。[17]

意大利蜂 
卡尼鄂拉蜂 
欧洲黑蜂 
高加索蜂 
 非洲化蜜蜂是西方蜜蜂欧洲亞種与非洲亞種(東非蜂)的雜交產物
非洲化蜜蜂是西方蜜蜂欧洲亞種与非洲亞種(東非蜂)的雜交產物
習性
生命周期
蜜蜂是完全变态的昆虫,发育周期包括卵(3天)、幼虫(6天)、蛹(12天)、成虫4个阶段。
- 蜂后将卵一个一个地产在蜂巢的每个巢室中。利用储精囊,蜂后可以决定要不要给所产的卵受精,通常取决于卵产在哪一种巢室内。蜂卵三天孵化成幼虫。
- 幼虫一开始会被年轻的工蜂(保育蜂)餵以蜂王漿,三天之后会换餵由蜂蜜和花粉酿造的蜂粮。幼虫再吃4天左右的蜂粮就停止饲餵,开始进入虫蛹期。那些被持续地、只餵以蜂王浆的幼虫,就会发育成蜂后。
- 幼虫会经历数次蜕皮,然后在巢室中吐丝编织出茧,把身子固定在蜂巢房里,再化成蛹。
- 成虫:

雄蜂的生殖器 - 年轻的工蜂(保育蜂)一开始负责清洁蜂巢与餵食幼虫。当分泌蜂王浆的腺体开始萎缩时,牠们开始负责建造蜂巢巢室。年龄更大的工蜂开始负责其他蜂群内部的任务,比如从採蜜蜂那里接收花蜜与花粉,以及保卫蜂巢。再到后来,工蜂才终于进行牠的第一次离巢飞行,成为一只採蜜蜂,直到牠死去。寿命在30–60天。
- 处女蜂后出房后五六天会远离母巢来到一个雄蜂聚集地进行交配飞行,牠会与数只雄蜂进行交配然后返回。婚飞后二三天开始产卵。如遇阴雨天,时间会推迟。雄蜂则在交配之后死去(寿命只有几个月)。蜂后不会与其母巢内的雄蜂进行交配。蜂后的寿命一般是3–5年,最长8–9年。
大多数蜜蜂王国并不是由单隻蜂后独立建立起来的,而是由一群蜜蜂——包括一只已交配的蜂后和一大支工蜂队伍——“分群”(swarming)而来。这群蜜蜂会全体迁移到一处筑巢地点,该地点已由工蜂事先侦查过,其方位也由特殊的舞蹈所告知。一旦这群蜜蜂到达那里,牠们会立即建造一个新的蜂巢,并开始哺育新的工蜂。这种建立巢穴的形式并没有在别的任何蜂属中发现,然而胡蜂科黄蜂的一些组(group)也是通过分群来建立新巢穴的(有时会有多只蜂后)。此外,无蛰蜂的新巢是由大量工蜂建立的,但新巢在蜂后入住之前就会建造完毕,这算不上真正的“分群”。

开放式蜂巢巢室内的蜜蜂卵 
蜂卵与幼虫 
蜜蜂蛹的发育阶段 .jpg.webp)
一只欧洲黑蜂破蛹而出
成虫形态

蜜蜂有1对复眼,较发达,位于头部两侧。由6000只小眼组成,表面布满细毛,可以分辨颜色和形状。雄蜂有3个单眼,位于额的上方,呈三角形排列;而蜂王和工蜂都只有一个单眼。雄蜂的复眼也比工蜂和蜂王的大,这样在牠和蜂王进行婚飞的时候可以更快地发现蜂王。一对触角位于两复眼之间的触角窝内,由柄节、梗节和鞭节构成,是最主要的触觉和嗅觉器官。
工蜂的嘴细长,雄蜂的嘴较短,而蜂王的嘴几乎看不到。蜜蜂的胸部有两双翅膀和三双腿,全身都张满了密密麻麻的短毛用来吸附花粉。
食物
蜜蜂的食物分为两个阶段,第一个阶段是从蜂卵孵化到变为幼虫的前3天,主要是吃王浆。3日龄以后,蜂王一直以蜂王浆为食,而工蜂和雄蜂主要吃蜂花粉和蜂蜜混成的蜂粮,直到蜜蜂幼虫封盖。蜜蜂变成成虫出房后主要吃蜂蜜。
蜜蜂所需的所有营养来自于多种多样的花粉和花蜜。花粉是自然界中蜜蜂的唯一蛋白质来源。成年的工蜂每天消耗3.4-4.3毫克的花粉来满足对干物质的需求(干物质中含66-74%的蛋白质)。[18]哺育一只幼虫需要125-187.5毫克的花粉或25-37.5毫克的蛋白质才能使其良好发育。[18]膳食蛋白质被分解为氨基酸,对蜜蜂来说10种氨基酸是必需的:蛋氨酸、色氨酸、精氨酸、赖氨酸、组氨酸、苯丙氨酸、异亮氨酸、苏氨酸、亮氨酸以及缬氨酸。其中所需浓度最高的是亮氨酸、异亮氨酸和缬氨酸,而精氨酸和赖氨酸的的高浓度是哺育幼蜂所需要的。[19]除了这些氨基酸以外,一些维他命B——包括生物素、叶酸、烟酰胺、核黄素、硫胺素、泛酸以及最重要的吡哆醇——也是哺育幼虫所需要的。吡哆醇是蜂王浆中最普遍存在的维他命B,浓度因觅食季节而异,五月份浓度最低,七八月份浓度最高。膳食吡哆醇缺失的蜜蜂是无法哺育幼蜂的。[19]
花粉也是蜜蜂的油脂来源。[18]油脂在哺育阶段被代谢掉,为之后的生物合成提供前体。脂溶性的维他命A、D、E和K虽然不是必需品,但却能显著地增加所哺育幼蜂的数量。[18]蜜蜂从花粉中消化植物甾醇以制造24-亚甲基胆甾醇和其他固醇,因为牠们无法从植物甾醇中直接合成胆固醇。哺育蜂能够选择性地在幼蜂的食物中添加固醇。[18]

蜜蜂採蜜 
蜜蜂採蜜 
牡丹和采食中的蜜蜂
花蜜被觅食的蜜蜂采集作为水分和碳水化合物(以蔗糖形式)的来源。蜜蜂食物中最主要的单糖是果糖和葡萄糖,但在血淋巴中最常见的循环糖类是海藻糖——一种包含两个葡萄糖分子的二糖。[20]成年工蜂每天需要摄入4毫克的可利用糖类,幼虫需要摄入约59.4毫克的碳水化合物才得以正常发育。[18]
蜜蜂需要摄入水来维持渗透压的动态平衡、制造液态的哺育食物,以及经蒸发作用为蜂巢降温。通过觅食花蜜,一个蜂群的水分需要大体都能得到满足,因为花蜜中水含量很高。在偶尔的炎热天气或者花蜜稀少的时候,觅食蜜蜂会从河流、池塘里取水以满足蜂巢的需要。[21]
| 蜂后 | 工蜂 | 雄蜂 | |
|---|---|---|---|
| 卵 | 受精卵 | 受精卵 | 未受精卵 |
| 性别 | ♀ 雌性 | ♀ 雌性 | ♂ 雄性 |
| 基因组 | 双倍体 | 双倍体 | 单倍体 |
| 卵房 | 王台 | 工蜂卵房 | 雄蜂卵房 |
| 食物 | 蜂王浆 | 蜂王浆/花粉花蜜 | 蜂王浆/花粉花蜜 |
| 发育时间(总计) | 16 天 | 21 天 | 24 天 |
| • 卵 | • 3 天 | • 3 天 | • 3 天 |
| • 幼虫 | • 5 天 | • 6 天 | • 7 天 |
| • 蛹 | • 8 天 | • 12 天 | • 14 天 |
| 重量 | 约200 mg | 约100 mg | 约200 mg |
| 身长 | 18–22 mm | 12–15 mm | 15–17 mm |
| 性成熟 | 约7 天 | - | 约14 天 |
| 寿命 | 3–4 年 | 4–7 月(冬季) 2–6 周(夏季) |
1–3 月 |
蜂鸣与振动
蜜蜂通过以间接飞行肌肉为主的胸部肌肉收缩,发生蜂鸣与振动。Mario Vallejo-Marín 建议将此分为三类。[22]
社会性
性别与阶级


一个蜜蜂群体有几千到几万只蜜蜂,由一只蜂后、少量的雄蜂和众多的工蜂组成[24]。工蜂和蜂后是从受精卵中孵化而来(雙倍體)。雄蜂是从未受精卵中孵化而来,只遗传了产卵蜂后的DNA,所以雄蜂的染色体数目都是正常数目的一半(單倍體)。产雄孤雌生殖方式——孤雌生殖的一个变种——控制着性别差异。性等位基因具有多态性,只要这对等位基因互不相同,则会产出雌蜂;如果这对等位基因相同,则会产出二倍体雄蜂。蜜蜂会侦查到二倍体雄蜂,并在孵化之后将牠们杀死。
- 蜂后是蜂群中唯一能正常产卵的雌性蜂。蜂后死後,蜂群会哺育新的蜂后。在蜂群的繁殖季节,蜂群会修筑多个王台哺育新的蜂后,一般老蜂后在新蜂后出台前会带领一部分蜜蜂离开原巢,并且第一个出台的新王会杀死未出台的新王继承老王的蜂巢成为新王。蜂后通常会在数次交配飞行中与数只雄蜂交配。一旦交配完成,蜂后将精子存贮在贮精囊中,开始产卵,并在需要时使用贮精囊中的精子进行受精。在某个特定地区,性等位基因的数目是限定的,因此,一只蜂后最有可能会与特定的一只或多只雄蜂交配——这些雄蜂的性等位基因与蜂后的其中一个性等位基因相同,于是蜂后通常会生产出一定比例的二倍体雄蜂卵。[lower-alpha 1]
- 雄蜂的体型要比工蜂大,全身呈黑色,腹部花纹不如工蜂明显。在蜂群中的作用是与蜂后交配,交配后立即死亡。雄蜂的精液可以在蜂后的体内保存数年而保持活力并具有授精能力。

- 工蜂是蜂群中繁殖器官发育不完善的雌性蜜蜂。在蜂群中为数众多,形态结构上表现出许多特化现象,如附肢特化形成花粉刷或花粉梳,生殖器官特化成尾端的螯刺,腹侧具腊腺。在同一蜂巢中的工蜂,因龄期不同可以分为三个生理上不同的工蜂群:保育蜂、筑巢蜂和採蜜蜂。保育蜂是刚羽化成的最年輕的工蜂,其主要职责是照管和饲养幼虫。当看管幼虫的工作要告一段落时,年輕的工蜂离巢短距离的飞翔,十天以后,蜡腺开始分泌蜂蜡,此时的工蜂称为筑巢蜂。筑巢蜂有时亦外出採蜜,或清扫蜂巢中的垃圾和死蜂。三周后,工蜂蜡腺停止分泌,成为专职的採蜜蜂。工蜂蜇人后,其蜇针連同腸臟 留在人体皮肤中,故其很快就会死亡。在蜜蜂群体中,数目最多,最为忙碌的可算是工蜂了。工蜂除了出外採蜜,供其他蜂群食用外,还有要负担保护蜂群不受袭击等工作,可是工蜂的地位非常低微,特别是连生育能力也没有。工蜂并非天生没有繁殖能力,而是在出生后被夺去。其实牠们可否发育成蜂后,这就是取决于牠们的出生后的饮食。从卵中孵化后,如果牠们连吃五天的蜂王浆,蜂体发育速度就快,16天就能发育成能生育的蜂后;但相反,若牠们只吃了两三天的蜂王浆,发育速度就会变得慢了,21天后才能变成虫,长大后牠们虽然仍是雌性的,但却失去了生育能力,成了工蜂。
- 工蜂会产卵,为不正常状态,因为都是未受精卵,而只能發育成雄蜂。
交流
工蜂之间互相协作以找到食物,牠们使用一种“舞蹈”的方式来互相交流关于资源的信息。这种舞蹈因物种而异,但蜜蜂属的所有物种在该行为上会展现出某种排列方式。如果资源距离蜂巢非常靠近,牠们可能也会展现出一种较普遍的舞蹈,通常称为“圆圈舞”。蜜蜂也会使用“鼓翼舞”来动员工蜂以接收采蜜蜂带回来的花粉花蜜。
调节温度
在寒冷地区,当气温降至约10 °C(50 °F)以下时,蜜蜂会停止飞行,牠们会挤进蜂巢的中心区域形成“过冬集群”。蜂后位于集群的中心,工蜂们簇拥在蜂后周围,靠身体不停颤动来使得中心温度保持在27 °C(81 °F)(冬季开始时的非产卵期)至34 °C(93 °F)(蜂后一旦恢复产卵)。工蜂们会轮流地从集群外围换到内部,使得每只蜜蜂都不至于太冷。集群最外边缘的温度会保持在大约8—9 °C(46—48 °F)。外界的气温越冷,集群会变得越紧密。在冬季期间,牠们会消耗所存储的蜂蜜来获得身体热量。冬季期间所消耗的蜂蜜数量取决于冬季长度和严寒程度,但在适中气候下大致范围会在15至50(33至110英磅)。[26]除此之外,某些蜜蜂——包括西方蜜蜂和东方蜜蜂——在冬夏时节气温改变时会採有一些有效机制来进行蜂巢的温度调节。夏日的温度调节方式是扇动翅膀以及从不同地方取水进行水分蒸发。[27]
防禦侵襲
蜜蜂的腹部末端長有長長的螫針,當人們因為害怕而撲打牠時,牠們就有可能出於自衛而螫人,螫人後牠們也會死去。這是因為螫針是由一根背刺針和兩根腹刺針組成,而螫針又與大、小毒腺和內臟器官相連,腹部刺針末端長有很多小倒鉤。當蜜蜂螫人時,小倒鉤會牢牢地勾緊人們的皮膚。當蜜蜂試圖飛離時,刺針就會連同部分內臟一起被拉出體外,蜜蜂也就因而死去。不過,當牠螫咬其牠有著硬質表皮的昆蟲時,因小倒鉤沒法勾緊光滑的表面,蜜蜂也就不會死去。雄蜂和蜂王不具备攻击能力,所以被蜜蜂螫伤的情形其实都是工蜂所为。
此外,蜜蜂不喜歡黑色物件,絨毛狀的東西,和酒,蔥,蒜等一類強烈的氣味。而女士常用的香水則會引來蜜蜂。
蜜蜂养殖
蜜蜂中的两个物种——西方蜜蜂和印度蜜蜂,经常被养蜂人所饲养和运输。现代的蜂房也使养蜂人得以运输蜜蜂,当作物需要授粉的时候将蜜蜂在田间转移,养蜂人则对授粉服务收取费用。这改变了长久以来个体经营的养蜂人的历史角色,也有利于大规模的商业运作。

产品

- 蜂蜜是蜜蜂对花蜜进行消化、处理之后的合成物,此物存贮在蜂巢中。[28]所有现存蜜蜂的蜂蜜都已被当地土著人采集以使用。西方蜜蜂和东方蜜蜂是仅有的两个被用于商业采蜜的物种。
- 巢蜜是完好地保存在巢脾中的食用蜂蜜,品质及价格要高于普通分离蜜[29]。
- 蜂花粉是花粉在蜂房内发酵后的产物[30]。
- 蜂蛹,一般是雄蜂蛹,因为个头大,而且雄蜂在蜂群中没有多大作用。
- 蜂蜡是长到特定年龄的工蜂从腹部蜡腺分泌的蜡[31]。
- 蜂胶是蜜蜂采集植物的汁液、花粉或花蜜,混合自己分泌的唾液与蜂蜡,所形成之胶状物。
- 蜂王浆是哺育工蜂(6–18日龄青年工蜂)的舌腺和上颚腺等腺体分泌的浆状物,为蜂王的终身食物及工蜂幼虫前三天的食物。




授粉服务
蜜蜂们都是花界全才,为许多开花植物授粉。在所有种类的蜜蜂中,只有西方蜜蜂被广泛用于商业上的水果与蔬菜作物的授粉。这些授粉服务的价值通常都在数十亿美元,在全世界作物价值的基础上增加了额外的约9%。蜜蜂的贡献基本上只在作物授粉领域,然而人们一直在讨论将牠的潜力推广到自然植被上,圈养蜜蜂与20000多种野生授粉者之间的竞争关系也同样饱受争议。[32]每年每个蜂房的蜜蜂能采集66英磅(30)的花粉。[33]
备注
- Work by Lechner in 2013 showed approximately 50 sex alleles in a population of Apis mellifera and estimated worldwide total number of sex alleles in A. mellifera between 116 and 145. There are likely more sex alleles in Apis cerana given wider distribution and larger number of geographical races. Honeybees are highly sensitive to inbreeding in part because duplication of the sex allele causes significant numbers of diploid drone eggs to be laid and subsequently destroyed as they hatch.[25]
參考文獻
- . 方舟馴服計算器. [2022-11-15]. (原始内容存档于2022-11-15).
- . pestworldforkids.org. [2016-04-26]. (原始内容存档于2021-05-10).
- . [2022-05-28]. (原始内容存档于2022-05-28).
- Douglas Harper. . 2006 [2016-02-27]. (原始内容存档于2017-07-06).
- Robert E. Snodgrass. . Cornell University Press. 1984: vii [2018-03-02]. ISBN 0-8014-9302-1. (原始内容存档于2021-04-14).
- . [February 26, 2016]. (原始内容存档于2021-03-18).
- . [February 21, 2016]. (原始内容存档于2021-04-17).
- . [2016-02-25]. (原始内容存档于2021-04-14).
- Deborah R. Smith; Lynn Villafuerte; Gard Otisc; Michael R. Palmer. (PDF). Apidologie. 2000, 31 (2): 265–279 [2018-03-02]. doi:10.1051/apido:2000121. (原始内容 (PDF)存档于2012-02-29).
- Michael S. Engel; I. A. Hinojosa-Diaz; A. P. Rasnitsyn. . Proceedings of the California Academy of Sciences. 2009, 60 (3): 23–38.
- Michael S. Engel. . Journal of Hymenoptera Research. 1999, 8: 165–196.
- Maria C. Arias; Walter S. Sheppard. . Molecular Phylogenetics and Evolution. 2005, 37 (1): 25–35. PMID 16182149. doi:10.1016/j.ympev.2005.02.017.Maria C. Arias; Walter S. Sheppard. . Molecular Phylogenetics and Evolution. 2005, 40 (1): 315. doi:10.1016/j.ympev.2006.02.002.
- Wongsiri, S., et al. "Comparative biology of Apis andreniformis and Apis florea in Thailand." Bee World 78.1 (1997): 23-35.
- Nathan Lo; Rosalyn S. Gloag; Denis L. Anderson; Benjamin P. Oldroyd. . Systematic Entomology. 2009, 35 (2): 226–233. doi:10.1111/j.1365-3113.2009.00504.x.
- . 知花蜂蜜网. 2016-03-26 [2020-12-07]. (原始内容存档于2022-06-11).
- . [2018-03-05]. (原始内容存档于2021-05-05).
- Head RJ. . The FARMS Review. 2008 [2018-03-05]. (原始内容存档于2013-07-30).
- Brodschneider, Robert; Crailsheim, Karl. . Apidologie. 2010-05-01, 41 (3): 278–294 [2018-03-09]. ISSN 0044-8435. doi:10.1051/apido/2010012. (原始内容存档于2021-02-24) (英语).
- Anderson, Leroy M; Dietz, A. . Apidologie. 1976. doi:10.1051/apido:19760105. (原始内容存档于2021-04-16).
- Karasov, William H.; Martinez del Rio, Carlos. . Princeton. 2008: 63–66.
- Kuhnholz, Susanne. . Behavioral Ecology and Sociobiology. 1997. doi:10.1007/s002650050402.
- Vallejo-Marín, Mario. . Journal of Experimental Botany. 2021-09-19, 73 (4). ISSN 0022-0957. PMC 8866655
 . PMID 34537837. doi:10.1093/jxb/erab428.
. PMID 34537837. doi:10.1093/jxb/erab428. - . . Harvard University Press. 2013-10-01: 525–584. ISBN 978-0-674-41851-6. doi:10.4159/harvard.9780674418516.c19 (英语).
- James L. Gould; Carol Grant Gould. . Scientific American Library. 1995: 19. ISBN 978-0-7167-6010-8.
- Lechner, Sarah; Ferretti, Luca; Schöning, Caspar; Kinuthia, Wanja; Willemsen, David; Hasselmann, Martin. . Molecular Biology and Evolution. 2014, 31 (2): 272–287. doi:10.1093/molbev/mst207.
- . [12 March 2016]. (原始内容存档于2016-03-04).
- Oldroyd, Benjamin P.; Wongsiri, Siriwat (2006). Asian Honey Bees (Biology, Conservation, and Human Interactions). Cambridge, Massachusetts and London, England: Harvard University Press.ISBN 0674021940.
- Crane E. . Ethology Ecology & Evolution. 1990, 3 (sup1): 100–105. doi:10.1080/03949370.1991.10721919.
- . 2010-03-17 [2011-12-01]. (原始内容存档于2012-03-09) (中文(简体)).
- Wong, Cathy. . about.com. February 2, 2005 [2011-04-10]. (原始内容存档于2012-06-03).
- Sanford, M.T.; Dietz, A. . Apidologie. 1976, 7: 197–207. doi:10.1051/apido:19760301.
- Geldmann, Jonas; González-Varo, Juan P. (PDF). Science. 2018, 359 (6374): 392–393 [2018-03-08]. doi:10.1126/science.aar2269. (原始内容存档 (PDF)于2018-02-09).
- . Back Yard Beekeepers Association. 2017 [2018-03-08]. (原始内容存档于2016-04-17).
- Bryony, Bonning. (PDF). Journal of Invertebrate Pathology. 11 November 2009, 103: s2-s4 [21 October 2014]. doi:10.1016/j.jip.2009.07.015.
- McDonald-Gibson, Charlotte. . The Independent. [2 July 2014]. (原始内容存档于2013-05-01).
- . Beeologics. [23 October 2014]. (原始内容存档于2014-11-07).
- Kwadha, Charles A.; Ong’amo, George O.; Ndegwa, Paul N.; Raina, Suresh K.; Fombong, Ayuka T. . Insects. 2017-06-09, 8 (2): 61 [2018-03-10]. doi:10.3390/insects8020061. (原始内容存档于2021-05-01) (英语).