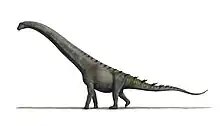
| Alamosaurus Temporal range: Late Cretaceous (Maastrichtian), | |
|---|---|
 | |
| Restored skeletons of Alamosaurus and Tyrannosaurus at Perot Museum | |
 | |
| Life reconstruction of Alamosaurus sanjuanensis | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Clade: | Saurischia |
| Clade: | †Sauropodomorpha |
| Clade: | †Sauropoda |
| Clade: | †Macronaria |
| Clade: | †Titanosauria |
| Family: | †Saltasauridae |
| Subfamily: | †Opisthocoelicaudiinae |
| Genus: | †Alamosaurus Gilmore, 1922 |
| Type species | |
| †Alamosaurus sanjuanensis Gilmore, 1922 | |
Alamosaurus (/ˌæləmoʊˈsɔːrəs/;[1] meaning "Ojo Alamo lizard") is a genus of opisthocoelicaudiine titanosaurian sauropod dinosaurs containing a single known species, Alamosaurus sanjuanensis, from the Maastrichtian age of the Late Cretaceous period in what is now southwestern North America. Isolated vertebrae and limb bones indicate that it reached sizes comparable to Argentinosaurus and Puertasaurus, which would make it the absolute largest dinosaur known from North America.[2] Its fossils have been recovered from a variety of rock formations spanning the Maastrichtian age. Specimens of a juvenile Alamosaurus sanjuanensis have been recovered from only a few meters below the Cretaceous-Paleogene boundary in Texas, making it among the last surviving non-avian dinosaur species.[3] Alamosaurus is the only known sauropod to have inhabited North America after their nearly 30-million year absence from the North American fossil record and probably represents an immigrant from South America.
Description
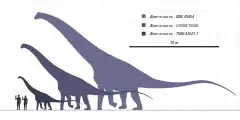
Alamosaurus was a gigantic quadrupedal herbivore with a long neck, long tail, and relatively long limbs.[3] Its body was at least partly covered in bony armor.[4] In 2012, Thomas Holtz gave a total length of 30 meters (98 ft) or more and an approximate weight of 72.5–80 tonnes (80–88 short tons) or more.[5][6] Though most of the complete remains come from juvenile or small adult specimens, three fragmentary specimens (SMP VP−1625, SMP VP−1850, and SMP VP−2104) suggest that adult Alamosaurus could have grown to enormous sizes comparable to the largest known dinosaurs, like Argentinosaurus, which has been estimated to weigh 73 metric tons (80 short tons).[2] Scott Hartman estimates Alamosaurus, based on a huge incomplete tibia that probably refers to it, being slightly shorter at 28–30 m (92–98 ft) and equal in weight to other massive titanosaurs, such as Argentinosaurus and Puertasaurus. It is currently the only titanosaur known from North America.[7] However, he says that, at the moment, scientists do not know whether the massive tibia belongs to an Alamosaurus or a completely new species of sauropod.[8]

In 2019, Gregory S. Paul estimated SMP VP−1625 at 27 tonnes (30 short tons) and he also mentioned a large partial anterior caudal vertebra that suggests an Alamosaurus specimen that is 15 percent dimensionally larger and with similar mass to his Dreadnoughtus estimation of 31 tonnes (34 short tons).[9] In 2020, Molina-Perez and Larramendi estimated the size of the largest individual at 26 meters (85.3 ft) and 38 tonnes (42 short tons).[10]

Though no skull has ever been found, rod-shaped teeth have been found with Alamosaurus skeletons and probably belonged to this dinosaur.[3][11] The vertebrae from the middle part of its tail had elongated centra.[12] Alamosaurus had vertebral lateral fossae that resembled shallow depressions.[12] Fossae that similarly resemble shallow depressions are known from Saltasaurus, Malawisaurus, Aeolosaurus, and Gondwanatitan.[12] Venenosaurus also had depression-like fossae, but its "depressions" penetrated deeper into the vertebrae, were divided into two chambers, and extend farther into the vertebral columns.[12] Alamosaurus had more robust radii than Venenosaurus.[12]
History

Alamosaurus remains have been discovered throughout the southwestern United States. The holotype was discovered in June of 1921 by Charles Whitney Gilmore, John Bernard Reeside,[13] and Charles Hazelius Sternberg at the Barrel Springs Arroyo in the Naashoibito Member of the Ojo Alamo Formation (or Kirtland Formation under a different definition) of New Mexico. This formation was deposited during the Maastrichtian age of the Late Cretaceous period.[14] Bones have also been recovered from other Maastrichtian formations, like the North Horn Formation of Utah and the Black Peaks, El Picacho, and Javelina Formations of Texas.[11] Undescribed titanosaur fossils closely associated with Alamosaurus have been found in the Evanston Formation in Wyoming. Three articulated caudal vertebrae were collected above Hams Fork and are housed at the Museum of Paleontology, University of California, Berkeley. However, these specimens have not been described.[15]

Smithsonian paleontologist Gilmore originally described holotype USNM 10486, a left scapula (shoulder bone), and the paratype USNM 10487, a right ischium (pelvic bone) in 1922, naming the type species Alamosaurus sanjuanensis. Contrary to popular assertions, the dinosaur is not named after the Alamo in San Antonio, Texas, or the battle that was fought there.[16] The holotype, the specimen the name was based on, was discovered in New Mexico and, at the time of its naming, Alamosaurus had not yet been found in Texas. Instead, the name Alamosaurus comes from Ojo Alamo, the geologic formation in which it was found and which was, in turn, named after the nearby Ojo Alamo trading post. Since this time, there has been some debate as to whether to reclassify the Alamosaurus-bearing rocks as belonging to the Kirtland Formation or if they should remain in the Ojo Alamo Formation. The term alamo itself is a Spanish word meaning "poplar" and is used for the local subspecies of cottonwood tree. The term saurus is derived from saura (σαυρα), the Greek word for "lizard", and is the most common suffix used in dinosaur names. There is only one species in the genus, Alamosaurus sanjuanensis, which is named after San Juan County, New Mexico, where the first remains were found.[14]
In 1946, Gilmore posthumously described a more complete specimen, USNM 15660, found on June 15, 1937, on the North Horn Mountain of Utah by George B. Pearce. It consists of a complete tail, a complete right forelimb (except for the fingers, which later research showed do not ossify with Titanosauridae), and both ischia.[17] Since then, hundreds of other bits and pieces from Texas, New Mexico, and Utah have been referred to Alamosaurus, often without much description. Despite being fragmentary, until the second half of the twentieth century they, represented much of the globally known titanosaurid material. The most completely known specimen, TMM 43621–1, is a juvenile skeleton from Texas which allowed educated estimates of length and mass.[3]
Some blocks catalogued under the same accession number as the relatively complete and well-known Alamosaurus specimen USNM 15660 and found in very close proximity to it based on bone impressions were first investigated by Michael Brett-Surman in 2009. In 2015, he reported that the blocks contained osteoderms, the first confirmation of their existence on Alamosaurus.[4]

The restored Alamosaurus skeletal mount at the Perot Museum [18] (pictured right) was discovered when student Dana Biasatti, a member of an excavation team at a nearby site, went on a hike to search for more dinosaur bones in the area.
Classification
In 1922, Gilmore was uncertain about the precise affinities of Alamosaurus and did not determine it any further than a general Sauropoda.[14] In 1927, Friedrich von Huene placed it in Titanosauridae.[19]
Alamosaurus was, in any case, an advanced and derived member of the group Titanosauria, but its relationships within that group are far from certain. The issue is further complicated by some researchers rejecting the name Titanosauridae and replacing it with Saltasauridae. One major analysis unites Alamosaurus with Opisthocoelicaudia in the subgroup Opisthocoelicaudiinae of Saltasauridae.[20] A major competing analysis finds Alamosaurus as a sister taxon to Pellegrinisaurus, with both genera located just outside Saltasauridae.[21] Other scientists have also noted particular similarities with the saltasaurid Neuquensaurus and the Brazilian Trigonosaurus (the "Peiropolis titanosaur"), which is used in many cladistic and morphologic analyses of titanosaurians.[3] A recent analysis published in 2016 by Anthony Fiorillo and Ron Tykoski indicates that Alamosaurus was a sister taxon to Lognkosauria and therefore to species such as Futalognkosaurus and Mendozasaurus, laying outside Saltasauridae (possibly being descended from close relations to the Saltasauridae), based on synapomorphies of cervical vertebral morphologies and two cladistic analyses.[22]The same study also suggests that the ancestors of Alamosaurus hailed from South America instead of Asia.[23]
Phylogeny
Alamosaurus in a cladogram after Navarro et al., 2022:[24]
| Saltasauridae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Age
Alamosaurus fossils are most notably found in the Naashoibito member of the Ojo Alamo Formation (dated to between about 69–68 million years old) and in the Javelina Formation, though the exact age range of the latter has been difficult to determine.[25] A juvenile specimen of Alamosaurus has been reported to come from the Black Peaks Formation, which overlies the Javelina in Big Bend, Texas, and also straddles the Cretaceous-Paleogene boundary. The Alamosaurus specimen was reported to come from a few meters below the boundary, dated to 66 million years ago, though the position of the boundary in this region is uncertain.[3] Only one geological site in the Javelina Formation has yielded the correct rock types for radiometric dating so far. The outcrop, situated in the middle strata of the formation about 90 meters (300 ft) below the K-Pg boundary and within the local range of Alamosaurus fossils, was dated to 69.0±0.9 million years old in 2010.[26] Using this date, in correlation with a measured age from the underlying Aguja Formation and the likely location of the K-Pg boundary in the overlying Black Peaks Formation, the Alamosaurus fauna seems to have lasted from about 70–66 million years ago, with the earliest records of Alamosaurus near the base of the Javelina formation and the latest just below the K-Pg boundary in the Black Peaks Formation.[26]
Biogeography
Alamosaurus is the only known sauropod to have lived in North America after the sauropod hiatus, a nearly 30-million-year interval for which no definite sauropod fossils are known from the continent. The earliest fossils of Alamosaurus date to the Maastrichtian age, around 70 million years ago, and it rapidly became the dominant large herbivore of southern Laramidia.[27]
The origins of Alamosaurus are highly controversial, with three hypotheses that have been proposed. The first of these, which has been termed the "austral immigrant" scenario,[28] proposes that Alamosaurus is descended from South American titanosaurs. Alamosaurus is closely related to South American titanosaurs, such as Pellegrinisaurus.[29][30] Alamosaurus appears in North America at the same time that hadrosaurs closely related to North American species first appear in South America, suggesting that the Alamosaurus lineage crossed into North America on the same routes as hadrosaurs crossed into South America.[31] The austral immigrant hypothesis has been challenged on the grounds that the routes connecting North and South America during the Maastrichtian may have consisted of separate islands, which would have presented challenges to the dispersal of titanosaurs.[27][32] A second scenario, termed the "inland herbivore" scenario,[28] suggests that titanosaurs were present in North America throughout the Late Cretaceous and that their apparent absence reflects the relative rarity of fossil sites preserving the upland environments that titanosaurs favored, rather than their true absence from the continent.[27] However, there is no evidence for sauropods in North America between the mid-Cenomanian and the early Maastrichtian, even in strata that preserve more upland environments, and the sauropods that lived in North America before the hiatus are basal titanosauriforms, such as Sonorasaurus and Sauroposeidon, not lithostrotian titanosaurs.[31][33] A third option is that, as in the austral immigrant scenario, Alamosaurus is not native to North America, but originated in Asia instead of South America.[32] Alamosaurus is commonly considered to be closely related to the Asian titanosaur Opisthocoelicaudia, but this is based on analyses that did not take Alamosaurus's South American relative Pellegrinisaurus into account.[29] Though many dinosaurs crossed between Asia and North America across the Bering land bridge, sauropods were poorly adapted for high-latitude environments and Beringia would have been an inhospitable environment for titanosaurs.[34] Furthermore, in order to reach southern Laramidia from Asia, Alamosaurus would have had to cross through Northern Laramidia, which contains no known sauropod fossils of comparable age to Alamosaurus, despite containing the best-studied dinosaur faunas on the continent.[22] Overall, a South American origin has been favored by several studies[30][22][29][34] and was regarded as "the only viable origin" for Alamosaurus by Chiarenza et al.[34]
Paleoecology
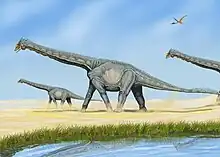
Skeletal elements of Alamosaurus are among the most common Late Cretaceous dinosaur fossils found in the United States Southwest and are now used to define the fauna of that time and place, known as the "Alamosaurus fauna". In the south of Late Cretaceous North America, the transition from the Edmontonian to the Lancian faunal stages is even more dramatic than it was in the north. Thomas M. Lehman describes it as "the abrupt reemergence of a fauna with a superficially 'Jurassic' aspect. These faunas are dominated by Alamosaurus and feature abundant Quetzalcoatlus in Texas. The Alamosaurus-Quetzalcoatlus association probably represent semi-arid inland plains.[27]
Contemporaries of Alamosaurus in the American southwest include unidentified tyrannosaurids similar to Tyrannosaurus, the oviraptorosaur Ojoraptorsaurus, the hadrosaurid Kritosaurus, the armored nodosaur Glyptodontopelta, and the chasmosaurine ceratopsids cf. Torosaurus utahensis, Bravoceratops, and Ojoceratops. Non-dinosaur taxa that had shared the same environment with Alamosaurus include the giant azhdarchid pterosaur Quetzalcoatlus, various species of fishes and rays, amphibians, lizards, turtles like Adocus, and multiple species of multituberculates like Cimexomys and Mesodma.
References
- ↑ "Alamosaurus". Lexico UK English Dictionary. Oxford University Press. Archived from the original on March 3, 2021.
- 1 2 Fowler, D. W.; Sullivan, R. M. (2011). "The First Giant Titanosaurian Sauropod from the Upper Cretaceous of North America". Acta Palaeontologica Polonica. 56 (4): 685. CiteSeerX 10.1.1.694.3759. doi:10.4202/app.2010.0105. S2CID 53126360.
- 1 2 3 4 5 6 Lehman, T.M.; Coulson, A.B. (2002). "A juvenile specimen of the sauropod Alamosaurus sanjuanensis from the Upper Cretaceous of Big Bend National Park, Texas" (PDF). Journal of Paleontology. 76 (1): 156–172. doi:10.1017/s0022336000017431. S2CID 232345559.
- 1 2 Carrano, M.T.; D'Emic, M.D. (2015). "Osteoderms of the titanosaur sauropod dinosaur Alamosaurus sanjuanensis Gilmore, 1922". Journal of Vertebrate Paleontology. 35 (1): e901334. doi:10.1080/02724634.2014.901334. S2CID 86797277.
- ↑ Holtz Jr., Thomas R. (2007). Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages. ISBN 978-0-375-82419-7. "Winter 2011 Apendix" (PDF).
- ↑ Holtz Jr., Thomas R. (2014). "Supplementary Information to Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages".
- ↑ "Assessing Alamosaurus". Skeletal Drawing.
- ↑ "The biggest of the big". Skeletal Drawing.
- ↑ Paul, Gregory S. (2019). "Determining the largest known land animal: A critical comparison of differing methods for restoring the volume and mass of extinct animals" (PDF). Annals of the Carnegie Museum. 85 (4): 335–358. doi:10.2992/007.085.0403. S2CID 210840060.
- ↑ Molina-Perez & Larramendi (2020). Dinosaur Facts and Figures: The Sauropods and Other Sauropodomorphs. New Jersey: Princeton University Press. p. 268. Bibcode:2020dffs.book.....M.
- 1 2 Weishampel, D.B. et al.. (2004). "Dinosaur Distribution (Late Cretaceous, North America)". In Weishampel, D.B., Dodson, P., Oslmolska, H. (eds.). "The Dinosauria (Second ed.)". University of California Press.
- 1 2 3 4 5 Tidwell, V., Carpenter, K. & Meyer, S. 2001. New Titanosauriform (Sauropoda) from the Poison Strip Member of the Cedar Mountain Formation (Lower Cretaceous), Utah. In: Mesozoic Vertebrate Life. D. H. Tanke & K. Carpenter (eds.). Indiana University Press, Eds. D.H. Tanke & K. Carpenter. Indiana University Press. 139–165.
- ↑ , John Bernard Reeside (1889-1958) was a geologist specializing in the study of the Mesozoic stratigraphy and paleontology of the western United States. While receiving his education at The Johns Hopkins University (A.B., 1911; Ph.D., 1915), he joined the United States Geological Survey (USGS) as a part-time assistant... [and] remained with the USGS for his entire professional career... From 1932 to 1949, Reeside was Chief of the Paleontology and Stratigraphy Branch.
- 1 2 3 Gilmore, C.W. (1922). "A new sauropod dinosaur from the Ojo Alamo Formation of New Mexico" (PDF). Smithsonian Miscellaneous Collections. 72 (14): 1–9.
- ↑ Lucas and Hunt, Spencer G. and Adrian P. (1989). Farlow, James O. (ed.). "Alamosaurus and the Sauropod Hiatus in the Cretaceous of the North American Western Interior". Paleobiology of the Dinosaurs. Geological Society of America Special Papers. 238 (238): 75–86. doi:10.1130/SPE238-p75. ISBN 0-8137-2238-1.
- ↑ Anthony D. Fredericks, 2012, Desert Dinosaurs: Discovering Prehistoric Sites in the American Southwest, The Countryman Press, p. 102-103
- ↑ Gilmore, C.W. 1946. Reptilian fauna of the North Horn Formation of central Utah. U.S. Geological Survey Professional Paper. 210-C:29–51.
- ↑ Perot Museum of Nature and Science
- ↑ v. Huene, F. (1927). "Sichtung der Grundlagen der jetzigen Kenntnis der Sauropoden". Eclogae Geologicae Helveticae. 20: 444–470.
- ↑ Wilson, J.A. (2002). "Sauropod dinosaur phylogeny: critique and cladistic analysis" (PDF). Zoological Journal of the Linnean Society. 136 (2): 217–276. doi:10.1046/j.1096-3642.2002.00029.x.
- ↑ Upchurch, P., Barrett, P.M. & Dodson, P. 2004. Sauropoda. In: Weishampel, D.B., Dodson, P., & Osmolska, H. (Eds.) The Dinosauria (2nd Edition). Berkeley: University of California Press. Pp. 259–322.
- 1 2 3 Tykoski, Ronald S.; Fiorillo, Anthony R. (2017). "An articulated cervical series of Alamosaurus sanjuanensis Gilmore, 1922 (Dinosauria, Sauropoda) from Texas: new perspective on the relationships of North America's last giant sauropod". Journal of Systematic Palaeontology. 15 (5): 1–26. doi:10.1080/14772019.2016.1183150.
- ↑ "Blogs". PLOS. Retrieved January 25, 2021.
- ↑ Navarro, Bruno A.; Ghilardi, Aline M.; Aureliano, Tito; Díaz, Verónica Díez; Bandeira, Kamila L. N.; Cattaruzzi, André G. S.; Iori, Fabiano V.; Martine, Ariel M.; Carvalho, Alberto B.; Anelli, Luiz E.; Fernandes, Marcelo A.; Zaher, Hussam (September 15, 2022). "A new nanoid titanosaur (Dinosauria: Sauropoda) from the Upper Cretaceous of Brazil". Ameghiniana. 59 (5): 317–354. doi:10.5710/AMGH.25.08.2022.3477. ISSN 1851-8044. S2CID 251875979.
- ↑ Sullivan, R.M., and Lucas, S.G. 2006. "The Kirtlandian land-vertebrate "age" – faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America." New Mexico Museum of Natural History and Science, Bulletin 35:7–29.
- 1 2 Lehman, T. M.; Mcdowell, F. W.; Connelly, J. N. (2006). "First isotopic (U-Pb) age for the Late Cretaceous Alamosaurus vertebrate fauna of West Texas, and its significance as a link between two faunal provinces". Journal of Vertebrate Paleontology. 26 (4): 922–928. doi:10.1671/0272-4634(2006)26[922:fiuaft]2.0.co;2. S2CID 130280606.
- 1 2 3 4 Lehman, Thomas M. (2001). "Late Cretaceous dinosaur provinciality". In Tanke, Darren H.; Carpenter, Kenneth (eds.). Mesozoic Vertebrate Life. Life of the past. Bloomington & Indianapolis: Indiana University Press. pp. 310–328. ISBN 0-253-33907-3.
- 1 2 <Lucas, Spencer G.; Hunt, Adrian P. (January 1, 1989). "Alamosaurus and the sauropod hiatus in the Cretaceous of the North American Western Interior". Paleobiology of the Dinosaurs. Geological Society of America Special Papers. 238: 75–86. doi:10.1130/SPE238-p75. ISBN 0-8137-2238-1.
- 1 2 3 Cerda, Ignacio; Zurriaguz, Virginia Laura; Carballido, José Luis; González, Romina; Salgado, Leonardo (July 21, 2021). "Osteology, paleohistology and phylogenetic relationships of Pellegrinisaurus powelli (Dinosauria: Sauropoda) from the Upper Cretaceous of Argentinean Patagonia". Cretaceous Research. 128: 104957. doi:10.1016/j.cretres.2021.104957. ISSN 0195-6671.
- 1 2 Gorscak, Eric; O‘Connor, Patrick M. (April 30, 2016). "Time-calibrated models support congruency between Cretaceous continental rifting and titanosaurian evolutionary history". Biology Letters. 12 (4): 20151047. doi:10.1098/rsbl.2015.1047. ISSN 1744-9561. PMC 4881341. PMID 27048465.
- 1 2 D'Emic, Michael D.; Wilson, Jeffrey A.; Thompson, Richard (2010). "The end of the sauropod dinosaur hiatus in North America". Palaeogeography, Palaeoclimatology, Palaeoecology. 297 (2): 486–490. Bibcode:2010PPP...297..486D. doi:10.1016/j.palaeo.2010.08.032. ISSN 0031-0182.
- 1 2 Mannion, Philip D.; Upchurch, Paul (January 15, 2011). "A re-evaluation of the 'mid-Cretaceous sauropod hiatus' and the impact of uneven sampling of the fossil record on patterns of regional dinosaur extinction". Palaeogeography, Palaeoclimatology, Palaeoecology. 299 (3): 529–540. Bibcode:2011PPP...299..529M. doi:10.1016/j.palaeo.2010.12.003. ISSN 0031-0182.
- ↑ D’Emic, Michael D.; Foreman, Brady Z. (July 2012). "The beginning of the sauropod dinosaur hiatus in North America: insights from the Lower Cretaceous Cloverly Formation of Wyoming". Journal of Vertebrate Paleontology. 32 (4): 883–902. doi:10.1080/02724634.2012.671204. ISSN 0272-4634. S2CID 128486488.
- 1 2 3 Chiarenza, Alfio Alessandro; Mannion, Philip D.; Farnsworth, Alex; Carrano, Matthew T.; Varela, Sara (December 17, 2021). "Climatic constraints on the biogeographic history of Mesozoic dinosaurs". Current Biology. 32 (3): 570–585.e3. doi:10.1016/j.cub.2021.11.061. hdl:11093/5013. ISSN 0960-9822. PMID 34921764. S2CID 245273901.