| C13orf42 | |||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||||||||||||||||||||||||||||||||||||||||||||
| Aliases | C13orf42, LINC00372, LINC00371, long intergenic non-protein coding RNA 371, chromosome 13 open reading frame 42 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| External IDs | GeneCards: C13orf42 | ||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||
| Wikidata | |||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||||


C13orf42 is a protein which, in humans, is encoded by the gene chromosome 13 open reading frame 42 (C13orf42). RNA sequencing data shows low expression of the C13orf42 gene in a variety of tissues.[3] The C13orf42 protein is predicted to be localized in the mitochondria, nucleus, and cytosol.[4] Tertiary structure predictions for C13orf42 indicate multiple alpha helices.[5]
Gene
Summary
C13orf42 is a protein encoding gene containing 4 exons. C13orf42 is also known by aliases LINC00371 and LINC00372.[3] RNA sequencing shows the gene's expression at low levels in various tissues.[3][6]
Location
C13orf42 is located on the minus strand of chromosome 13 at 13q14.3 in humans.[3][7] C13orf42 is located from 51.08 Mb to 51.20 Mb on chromosome 13 and spans 118 kilobases.[8]
Neighborhood
The genomic neighborhood of C13orf42 consists of several pseudogenes along with ribonuclease H2 subunit B (RNASEH2B), uncharacterized LOC107984554, and family with sequence similarity 124 member A (FAM124A).[3]
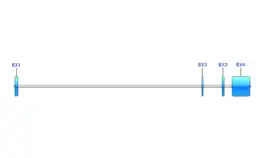
Exons
Expression
RNA sequencing of C13orf42 shows expression in a variety of tissues including the spleen, kidney, heart, brain, testis, skin, esophagus, colon, small intestine, stomach, lung, placenta, salivary gland, thymus, and adipose.[3] RNA sequencing of human fetal tissue shows C13orf42 expression starting at 20 weeks in the intestine, 16 weeks in the kidney, 10 weeks in the lung, and expression in the stomach is seen at 16 weeks but not 10, 18, or 20 weeks.[3] Recorded RNA expression is very low, with all results being lower than 0.5 reads per kilobase of transcript per million reads mapped (RPKM). Microarray data from NCBI geo (GDS425) shows expression in additional tissues including bone marrow, liver, skeletal muscle, spinal cord, and pancreas.[6]
Transcript
Variants
C13orf42 produces four known transcript variants, variant 1, variant 2, variant 3, and variant X1. Transcript variant 3 (accession number: NM_001351589.3) is the longest high-quality mRNA at 3075 nucleotides.[10] Transcript variant 3 contains 4 exons and encodes a 325 amino acid protein.
Transcript variants 1, 2, and X1 all lack the first exon but align with exons 2, 3, and 4 of transcript variant 3. Variants 1 and 2 are not protein encoding, while variants 3 and X1 are protein coding. Variant X1 is 2717 nucleotides long and encodes a 189 amino acid protein which aligns with the last 187 amino acids of the longer protein encoded by transcript variant 3 and differs in its first two amino acids.[11][12]
Protein
Isoforms
There are two known proteins encoded by the isoforms of C13orf42.[13] Transcript variant 3 encodes the longest protein at 325 amino acids long.[13] Transcript variant X1 encodes a 189 amino acid long protein.[13] This protein aligns with exons 2, 3, and 4 of the 325 amino acid protein, but is missing exon 1.[12]
Protein Composition
C13orf42 has a predicted isoelectric point of 9.3 and a predicted molecular weight of 37.4 kDa.[14] Human C13orf42 is a serine rich and positively charged amino acid (lysine and arginine) rich protein.[15] This composition is partially conserved in orthologs.
Tertiary Structure
The C13orf42 tertiary structure of the highest confidence predicted by I-Tasser is predicted to have many alpha helices.[5] In the structure below, residues indicated to be present in C13orf42 in higher amounts (serine, lysine and arginine) are annotated.[16] A space filling model and a charge model is also shown for C13orf42.
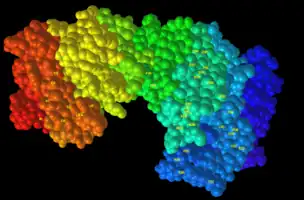

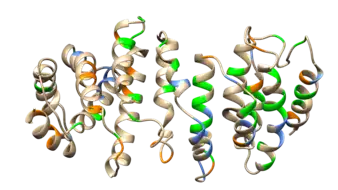
Subcellular Localization
Human C13orf42 is predicted to be localized to the mitochondria, nucleus, cytosol, and endoplasmic reticulum with the ER predicted at a low percentage (<5%).[4] Orthologs show similar predicted subcellular localization with mitochondria, nucleus, and cytosol being the top predicted locations, however, predicted percentages vary.[4]
Immunohistochemistry
C13orf42 antibody B-4 (catalog number: sc-376095) shows cytoplasmic and nuclear staining in seminiferous ducts and Lyedig cells of testis tissue.[18] C13orf42 antibody E-3 (catalog number: sc-374567) shows cytoplasmic staining in seminiferous ducts and Lyedig cells of testis tissue, and cytoplasmic and nucleolar localization in HeLa cells.[19]
Post translational Modifications
C13orf42 is predicted to have 10 highly conserved (in over 70% of analyzed orthologs from table below) phosphorylation sites.[20] Phosphorylation sites include one CK2 phosphorylation, one TYR phosphorylation, two cAMP phosphorylation sites, and six PKC phosphorylation sites. There are three predicted O-β-GlcNAc sites and two predicted yin-yang sites in C13orf42 which are fully conserved in orthologs.[21] A yin-yang site occurs when O-β-GlcNAc and phosphorylation are predicted for the same site. C13orf42 is not predicted to have myristylation sites as it does not contain an N-terminal glycine.[22]
Domains
C13orf42 has no identified domains with high confidence or conservation in orthologs.
Homology and evolution

Orthologs
C13orf42 has orthologs in mammals, birds, reptiles, amphibians, bony fish, and cartilaginous fish as shown in the ortholog table below.[23] No orthologs were found in jawless fish, invertebrates, plants, fungi, viruses, or bacteria.[23] All mammals contain the same 4 exons as the human C13orf42 protein, and nonmammals are missing exon 4. Mammalian orthologs have a high percent identity to human C13orf42, each having over 62% identity. The furthest orthologs (cartilaginous fish) have sequence identities around 33%. Human C13orf42 does not have paralogs.[8]
Ortholog Table
| Genus and Species | Common Name | Taxonomic Class | Date of Divergence (MYA)[24] | Accession Number | Length (amino acids) | Percent Identity to Homo sapiens | Percent Similarity to Homo sapiens |
|---|---|---|---|---|---|---|---|
| Homo sapiens | Human | Primates | 0 | NP_001338518.1 | 325 | 100 | 100 |
| Mus musculus | Mouse | Dasyuromorphia | 87 | XP_030104110.1 | 318 | 76.9 | 84 |
| Tursiops truncatus | Common bottlenose dolphin | Cetacea | 94 | XP_033699576.1 | 319 | 82.8 | 87.7 |
| Equus caballus | Horse | Perissodactyla | 94 | XP_023477317.1 | 326 | 82.6 | 90.2 |
| Mustela putorius | European polecat | Carnivora | 94 | XP_004775284.1 | 326 | 79.4 | 85 |
| Pipistrellus kuhlii | Kuhl's pipistrelle (bat) | Chiroptera | 94 | XP_036312978.1 | 325 | 79.1 | 86.8 |
| Ursus maritimus | Polar bear | Ursidae | 94 | XP_040478472.1 | 328 | 77.2 | 82.7 |
| Elephas maximus indicus | Indian elephant | Proboscidea | 99 | XP_049709344.1 | 326 | 79.4 | 86.2 |
| Dasypus novemcinctus | Nine-banded armadillo | Cingulata | 99 | XP_023446856.1 | 328 | 72.8 | 81.4 |
| Dromiciops gliroides | Monito del monte | Microbiotheria | 160 | XP_043849658.1 | 326 | 67.9 | 79.5 |
| Trichosurus vulpecula | Common brushtail possum | Diprotodontia | 160 | XP_036599801.1 | 326 | 66.7 | 79.5 |
| Sarcophilus harrisii | Tasmanian devil | Dasyuromorphia | 160 | XP_023355407.1 | 326 | 66.4 | 80.1 |
| Ornithorhynchus anatinus | Playtpus | Monotremes | 180 | XP_028904285.1 | 330 | 62.3 | 73 |
| Alligator sinensis | Chinese alligator | Crocodilian | 319 | XP_025062978.1 | 266 | 48.6 | 61.7 |
| Dromaius novaehollandiae | Emu | Casuariiformes | 319 | XP_025964173.1 | 268 | 48.2 | 59.8 |
| Gallus gallus | Chicken | Galliformes | 319 | XP_004938779.1 | 268 | 47.6 | 61.3 |
| Camarhynchus parvulus | Small tree finch | Thraupidae | 319 | XP_030802909.1 | 264 | 47.1 | 60.9 |
| Pelodiscus sinensis | Chinese softshell turtle | Testudines | 319 | XP_014429996.1 | 267 | 47 | 60.6 |
| Phasianus colchicus | Common pheasant | Galliformes | 319 | XP_031471701.1 | 272 | 46.4 | 59.9 |
| Varanus komodoensis | Komodo dragon | Squamata | 319 | XP_044300138.1 | 269 | 45.9 | 60.1 |
| Python bivittatus | Burmese python | Squamata | 319 | XP_025026122.1 | 269 | 44.1 | 58 |
| Pantherophis guttatus | Corn snake | Squamata | 319 | XP_034287884.1 | 267 | 42.7 | 56.4 |
| Rhinatrema bivittatum | Two-lined caecilian | Rhinatrematidae | 353 | XP_029459031.1 | 273 | 45.4 | 59.5 |
| Xenopus tropicalis | Western clawed frog | Anura | 353 | XP_031752228.1 | 260 | 43.5 | 57.4 |
| Bufo gargarizans | Asiatic toad | Anura | 353 | XP_044141363.1 | 262 | 40.7 | 56.8 |
| Bufo bufo | Common toad | Anura | 353 | XP_040278366.1 | 391 | 30.9 | 42 |
| Protopterus annectens | West african lungfish | Lepidosireniformes | 408 | XP_043927617.1 | 269 | 37.8 | 53.5 |
| Polyodon spathula | Paddlefish | Acipenseriformes | 431 | XP_041127510.1 | 275 | 33.8 | 52.4 |
| Danio rerio | Zebrafish | Cypriniformes | 431 | XP_021329868.1 | 287 | 30.2 | 44.7 |
| Clupea harengus | Atlantic herring | Clupeiformes | 431 | XP_042559186.1 | 306 | 29 | 41.7 |
| Callorhinchus milii | Australian ghostshark | Chimaeriformes | 464 | XP_042189981.1 | 267 | 35.5 | 53.3 |
| Amblyraja radiata | Thorny skate | Rajiformes | 464 | XP_032889397.1 | 267 | 33 | 47.5 |
| Scyliorhinus canicula | Small-spotted catshark | Carcharhiniformes | 464 | XP_038658253.1 | 264 | 32.5 | 50.4 |
Phylogeny
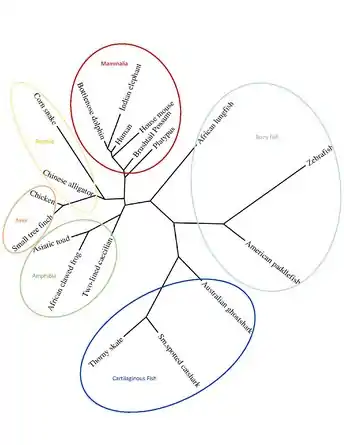
A phylogenetic tree shows human C13orf42 is most related its mammalian orthologs, and most distantly related to cartilaginous fish orthologs.[25]
Function
Clinical significance
Kanagal-Shamanna et. al identified an ATM fusion with C13orf42 in a patient with chronic lymphocytic leukemia which lead to ATM inactivation.[26]
Xiong et. al indicated SNP rs7325564 to be significantly associated with nasion and pronasale face shape in humans.[27]
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000226792 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- 1 2 3 4 5 6 7 "C13orf42 chromosome 13 open reading frame 42 [Homo sapiens (human)] - Gene - NCBI". www.ncbi.nlm.nih.gov.
- 1 2 3 "PSORT II Prediction". psort.hgc.jp.
- 1 2 3 4 5 Yang J, Zhang Y (July 2015). "I-TASSER server: new development for protein structure and function predictions". Nucleic Acids Research. 43 (W1): W174–W181. doi:10.1093/nar/gkv342. PMC 4489253. PMID 25883148.
- 1 2 Yanai I, Benjamin H, Shmoish M, Chalifa-Caspi V, Shklar M, Ophir R, et al. (March 2005). "Genome-wide midrange transcription profiles reveal expression level relationships in human tissue specification". Bioinformatics. 21 (5): 650–659. doi:10.1093/bioinformatics/bti042. PMID 15388519.
- ↑ "C13orf42 Gene - GeneCards | CM042 Protein | CM042 Antibody". www.genecards.org.
- 1 2 3 Kent WJ, Sugnet CW, Furey TS, Roskin KM, Pringle TH, Zahler AM, Haussler D (June 2002). "The human genome browser at UCSC". Genome Research. 12 (6): 996–1006. doi:10.1101/gr.229102. PMC 186604. PMID 12045153.
- 1 2 Liu W, Xie Y, Ma J, Luo X, Nie P, Zuo Z, et al. (October 2015). "IBS: an illustrator for the presentation and visualization of biological sequences". Bioinformatics. 31 (20): 3359–3361. doi:10.1093/bioinformatics/btv362. PMC 4595897. PMID 26069263.
- 1 2 "Homo sapiens chromosome 13 open reading frame 42 (C13orf42), transcript variant 3, mRNA". NCBI nucleotide. 11 June 2022.
- ↑ "PREDICTED: Homo sapiens chromosome 13 open reading frame 42 (C13orf42), transcript variant X1, mRNA". 5 April 2022.
- 1 2 3 Needleman SB, Wunsch CD (March 1970). "A general method applicable to the search for similarities in the amino acid sequence of two proteins". Journal of Molecular Biology. 48 (3): 443–53. doi:10.1016/0022-2836(70)90057-4. PMID 5420325.
- 1 2 3 4 5 "Home - Protein - NCBI". www.ncbi.nlm.nih.gov.
- ↑ Wilkins MR, Gasteiger E, Bairoch A, Sanchez JC, Williams KL, Appel RD, Hochstrasser DF (24 September 1998). "Protein identification and analysis tools in the ExPASy server". 2-D Proteome Analysis Protocols. Methods in Molecular Biology. Vol. 112. pp. 531–552. doi:10.1385/1-59259-584-7:531. ISBN 1-59259-584-7. PMID 10027275.
- 1 2 Brendel V, Bucher P, Nourbakhsh IR, Blaisdell BE, Karlin S (March 1992). "Methods and algorithms for statistical analysis of protein sequences". Proceedings of the National Academy of Sciences of the United States of America. 89 (6): 2002–2006. Bibcode:1992PNAS...89.2002B. doi:10.1073/pnas.89.6.2002. PMC 48584. PMID 1549558.
- 1 2 Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, Ferrin TE (October 2004). "UCSF Chimera--a visualization system for exploratory research and analysis". Journal of Computational Chemistry. 25 (13): 1605–1612. doi:10.1002/jcc.20084. PMID 15264254. S2CID 8747218.
- 1 2 Wang J, Youkharibache P, Zhang D, Lanczycki CJ, Geer RC, Madej T, et al. (January 2020). "iCn3D, a web-based 3D viewer for sharing 1D/2D/3D representations of biomolecular structures". Bioinformatics. 36 (1): 131–135. doi:10.1093/bioinformatics/btz502. PMC 6956771. PMID 31218344.
- ↑ "C13orf42 Antibody (B-4)". www.scbt.com.
- ↑ "C13orf42 Antibody (E-3)". www.scbt.com.
- ↑ "Motif Scan". myhits.sib.swiss.
- ↑ Blom N, Sicheritz-Pontén T, Gupta R, Gammeltoft S, Brunak S (June 2004). "Prediction of post-translational glycosylation and phosphorylation of proteins from the amino acid sequence". Proteomics. 4 (6): 1633–1649. doi:10.1002/pmic.200300771. PMID 15174133. S2CID 18810164.
- ↑ Bologna G, Yvon C, Duvaud S, Veuthey AL (June 2004). "N-Terminal myristoylation predictions by ensembles of neural networks". Proteomics. 4 (6): 1626–1632. doi:10.1002/pmic.200300783. PMID 15174132. S2CID 20289352.
- 1 2 "C13orf42 BLAST: Basic Local Alignment Search Tool". blast.ncbi.nlm.nih.gov.
- ↑ Kumar S, Suleski M, Craig JM, Kasprowicz AE, Sanderford M, Li M, et al. (August 2022). "TimeTree 5: An Expanded Resource for Species Divergence Times". Molecular Biology and Evolution. 39 (8): msac174. doi:10.1093/molbev/msac174. PMC 9400175. PMID 35932227.
- 1 2 Dereeper A, Guignon V, Blanc G, Audic S, Buffet S, Chevenet F, et al. (July 2008). "Phylogeny.fr: robust phylogenetic analysis for the non-specialist". Nucleic Acids Research. 36 (Web Server issue): W465–W469. doi:10.1093/nar/gkn180. PMC 2447785. PMID 18424797.
- ↑ Kanagal-Shamanna R, Bao H, Kearney H, Smoley S, Tang Z, Luthra R, et al. (April 2022). "Molecular characterization of Novel ATM fusions in chronic lymphocytic leukemia and T-cell prolymphocytic leukemia". Leukemia & Lymphoma. 63 (4): 865–875. doi:10.1080/10428194.2021.2010061. PMID 34898335. S2CID 245131979.
- ↑ Xiong Z, Dankova G, Howe LJ, Lee MK, Hysi PG, de Jong MA, et al. (November 2019). McCarthy MI, Morris AP, Cha S (eds.). "Novel genetic loci affecting facial shape variation in humans". eLife. 8: e49898. doi:10.7554/eLife.49898. PMC 6905649. PMID 31763980.