| Dacrytherium Temporal range: Middle Eocene to late Eocene | |
|---|---|
 | |
| Dacrytherium ovinum mandibles, National Museum of Natural History, France | |
| Scientific classification | |
| Domain: | Eukaryota |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Artiodactyla |
| Family: | †Anoplotheriidae |
| Subfamily: | †Dacrytheriinae |
| Genus: | †Dacrytherium Filhol, 1876 |
| Type species | |
| †Dichobune ovina (= †Dacrytherium ovinum) Owen, 1857 | |
| Other species | |
| |
| Synonyms | |
|
Genus synonymy
Synonyms of D. ovinum
| |
Dacrytherium (Ancient Greek: δάκρυ (tear) + θήρ (beast or wild animal) meaning "tear beast") is an extinct genus of Paleogene artiodactyls belonging to the family Anoplotheriidae. It occurred from the middle to late Eocene of western Europe and is the type genus of the subfamily Dacrytheriinae, the older of the two anoplotheriid subfamilies. Dacrytherium was first erected in 1876 by the French palaeontologist Henri Filhol, who recognized in his studies that it had dentition similar to the anoplotheriids Anoplotherium and Diplobune but differed from them by a deep preorbital fossa, where the genus name derives from. Since then, there are currently four valid species within the genus, of which D. ovinum is the type species.
Dacrytherium was a medium-sized artiodactyl that is defined by specific dental traits separating it from the Anoplotheriinae. Typically, its species had a deep preorbital fossa that anoplotheriines lack entirely, although D. elegans had a different morphology for the depression. Its dental and cranial morphologies, however, were otherwise typical of the Anoplotheriidae, which led to historic confusions regarding whether Dacrytherium belonged to the Anoplotheriidae or its own family. It is also recognized as having two lineages in the form of D. elegans-D. saturnini and D. priscum-D. ovinum given the morphology changes and size increases of dentitions, although the former lineage has been doubted.
Dacrytherium lived in western Europe during a period when the region was an archipelago isolated from the rest of Eurasia. Contemporary species from this region were not widely dispersed due to high levels of endemism. It co-existed with a wide variety of artiodactyls and perissodactyls in subtropical-tropical environments that supported frugivorous-folivorous mammals. Dacrytherium itself was likely folivorous, but its behaviours are unknown given the general scarcity of post-cranial evidence and the unusual variations of hypothesized behaviours in the derived anoplotheriines Anoplotherium and Diplobune.
Taxonomy
Early history
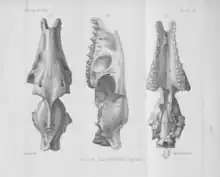
In 1876, French palaeontologist Henri Filhol described fossils from recent excavations at the phosphorite deposits of Quercy in France, including bones that he identified as belonging to new genera or species. One "pachyderm", he said, had a dental formula, in terms of the incisors, canines, and molars, of 3.1.73.1.7, which he considered similar in character to other fossil mammal genera like Anthracotherium. The specimen that he had was a complete skull with a "peculiar" upper jaw and a lower jaw and all of its teeth. From the skull, he erected the genus name Dacrytherium and established the species D. anthracoides.[1] The same year, Paul Gervais referenced Dacrytherium, previously erected by Filhol, calling it "interesting" based on the upper maxilla of the complete skull having a deep lateral (or outer) hollowing similar to the hipparionines or merycoidodonts.[2] The etymology of the genus name derives in Greek from "dacry(o)" (tear) and "thḗr" (beast or wild animal) meaning "tear beast", referencing the lacrimal fossa.[3]
In 1877, however, Filhol retained the genus name but replaced the species name "D. anthracoides" with "D. cayluxi" while displaying fossil evidence in his source.[4][5] He emphasized the upper complete skull of Dacrytherium that he found from the Lamandine-Haute deposits in 1876, which had a lower jaw and some leg bones that potentially belonged to it. Filhol determined that its dental formula was 3.1.4.33.1.4.3. The dentition, he said, was more similar to Anoplotherium and Diplobune than Xiphodon.[5]
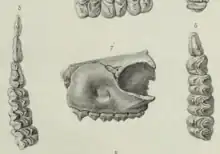
Filhol described another small species of "pachyderm" from the phosphorites of Lamandine-Haute in 1884 based on a fragment of a skull with premolars and molars. He stated that it was very similar to Anoplotherium, that the first and second premolars were "almost absolutely identical" to those of Dacrytherium. Based on very specific molar differences, however, he proposed the genus and species name Plesidacrytherium elegans. Like Dacrytherium, the palaeontologist said, P. elegans presented a depression at the area of the infraorbital foramen up the upper maxilla.[6] Filhol already created the name Plesydacrytherium elegans as early as 1880,[7] but it lacked any actual definition to make it valid.[8] The genus name derived in Ancient Greek from "plēsíon" (near), "dacry(o)" (tear), and "thḗr" (beast or wild animal) meaning "near tear beast".[3]
British naturalist Richard Lydekker reviewed known species of anoplotheriids in 1885. Previously in 1857, Richard Owen described an "anoplotheroid quadruped" the size of Xiphodon gracilis from the Isle of Wight. Owen then determined that because of its dentition corresponding more to Dichobune than to Xiphodon and Dichodon, the specimens belonged to the newly erected Dichobune ovina. Lydekker determined that the so-called "Dichobune ovinus" actually belonged to Dacrytherium. He then said that Dacrytherium ovinum was a larger form of D. cayluxi (or D. cayluxense). In addition, he considered that "Xiphodon platyceps" may be the same as D. ovinum.[9][10]
Later revisions

In 1891-1893, Karl Alfred von Zittel synonymized Mixtotherium plus Plesidacrytherium with Diplobune and Adrotherium (species A. depressum) with Dacrytherium. He supported the validity of D. cayluxense and D. ovinum as well as the synonymy of "X. platyceps" with the latter species.[11] However, in 1892, Lydekker made reviews of the species of Dacrytherium. He said that he previously reclassified "Dichobune ovina" to Dacrytherium based on the dental series of a cranium. At that time, he thought that D. cayluxense was one of two distinct species of Dacrytherium because he thought that the former differed from the latter based on dental differences including the canines. Just a few months before Lydekker wrote his 1892 article, Arthur Smith Woodward presented him with more fossil evidence from the French phosphorites including a well-preserved mandible of D. cayluxi, leading him to synonymize it with D. ovinum. In addition, Lydekker considered his previous synonymy of X. playceps with Dacrytherium "unfounded" since the former clearly lacked any lacrimal fossa that the latter has.[12]
In 1908, Hans Georg Stehlin synonymized Adrotherium with Mixtotherium instead of Dacrytherium.[13] Also in 1910, Stehlin reaffirmed the validity of Mixtotherium and synonymized Plesidacrytherium with Dacrytherium, reclassifying its species to D. elegans. He synonymized "X. platyceps" with Dichodon cuspidatum because of their close dentitions combined with the former lacking the preorbital fossa that Dacrytherium has. In his revisions of the Paleogene artiodactyls, he erected two more species of Dacrytherium: D. priscum and D. saturnini. The name D. priscum refers to a larger form of the genus. According to Stehlin, Ludwig Rütimeyer knew of specimens of Dacrytherium in 1891 but doubted that it actually belonged to the genus based on Lydekker's specimen illustrations, a conclusion that Stehlin considered a mistake. The second species D. saturnini was also based on specific cranial plus dental differences and was named after the locality of Saint-Saturnin-lès-Apt in the department of Vaucluse, France where the fossils came from.[14][15]
Classification
.jpg.webp)
Dacrytherium is the type genus of the subfamily Dacrytheriinae, which belongs to the Paleogene artiodactyl family Anoplotheriidae.[16] The family was endemic to western Europe and lived from the middle Eocene to the early Oligocene (~44 to 30 Ma, possible earliest record at ~48 Ma). The exact evolutionary origins and dispersals of the anoplotheriids are uncertain, but they exclusively resided within the continent when it was an archipelago that was isolated by seaway barriers from other regions such as Balkanatolia and the rest of eastern Eurasia. The Anoplotheriidae's relations with other members of the Artiodactyla are not well-resolved, with some determining it to be either a tylopod (which includes camelids and merycoidodonts of the Paleogene) or a close relative to the infraorder and some others believing that it may have been closer to the Ruminantia (which includes tragulids and other close Paleogene relatives).[17][18]
The history of dacrytheriines has been contentious as a result of disagreements as to whether they constitute a subfamily of the Anoplotheriidae or a distinct family named "Dacrytheriidae". The family name was first proposed by Charles Depéret in 1917 and was generally followed upon for decades by other palaeontologists like Jean Sudre. Since 2007, however, they were redefined as a subfamily within the Anoplotheriidae, supported by recent phylogenetic analyses determining that Dacrytherium falls within the Anoplotheriidae. It is one of two subfamilies of the Anoplotheriidae, the other being the Anoplotheriinae.[16][19][20]
The Dacrytheriinae is the older anoplotheriid subfamily, but the actual first appearance by Mammal Paleogene zone range is uncertain. The first undisputed appearance of anoplotheriids is by MP13, but their range may have extended, in the case of Catodontherium, into MP11 or even MP10.[16][21] Dacrytherium itself made its first undisputed appearance by MP13 as an artiodactyl leaning towards bunoselenodont dentition.[22] The younger subfamily Anoplotheriinae made their first appearances by the late Eocene (MP15-MP16), or ~41-40 Ma, within western Europe with Duerotherium and Robiatherium. After a significant gap of anoplotheriines in MP17a-MP17b, the derived anoplotheriids Anoplotherium and Diplobune made their first appearances in western Europe by MP18, although their exact origins are unknown.[23] The Dacrytheriinae has recently been suggested to have been a paraphyletic subfamily based on dental morphology from which the Anoplotheriinae, Mixtotheriidae, and Cainotherioidea stemmed, but further research is required to confirm if this is true.[24]
Conducting studies focused on the phylogenetic relations within the Anoplotheriidae has proven difficult due to the general scarcity of fossil specimens of most genera.[23] The phylogenetic relations of the Anoplotheriidae as well as the Xiphodontidae, Mixtotheriidae, and Cainotheriidae have also been elusive due to the selenodont morphologies of the molars, which were convergent with tylopods or ruminants.[24] Some researchers considered the selenodont families Anoplotheriidae, Xiphodontidae, and Cainotheriidae to be within Tylopoda due to postcranial features that were similar to the tylopods from North America in the Paleogene.[25] Other researchers tie them as being more closely related to ruminants than tylopods based on dental morphology. Different phylogenetic analyses have produced different results for the "derived" selenodont Eocene European artiodactyl families, making it uncertain whether they were closer to the Tylopoda or Ruminantia.[20][26]
In an article published in 2019, Romain Weppe et al. conducted a phylogenetic analysis on the Cainotherioidea within the Artiodactyla based on mandibular and dental characteristics, specifically in terms of relationships with artiodactyls of the Paleogene. The results retrieved that the superfamily was closely related to the Mixtotheriidae and Anoplotheriidae. They determined that the Cainotheriidae, Robiacinidae, Anoplotheriidae, and Mixtotheriidae formed a clade that was the sister group to the Ruminantia while Tylopoda, along with the Amphimerycidae and Xiphodontidae split earlier in the tree.[26] The phylogenetic tree used for the journal and another published work about the cainotherioids is outlined below:[19]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
In 2022, Weppe created a phylogenetic analysis in his academic thesis regarding Paleogene artiodactyl lineages, focusing most specifically on the endemic European families. The phylogenetic tree, according to Weppe, is the first to conduct phylogenetic affinities of all anoplotheriid genera, although not all individual species were included. He found that the Anoplotheriidae, Mixtotheriidae, and Cainotherioidea form a clade based on synapomorphic dental traits (traits thought to have originated from their most recent common ancestor). The result, Weppe mentioned, matches up with previous phylogenetic analyses on the Cainotherioidea with other endemic European Paleogene artiodactyls that support the families as a clade. As a result, he argued that the proposed superfamily Anoplotherioidea, composing of the Anoplotheriidae and Xiphodontidae as proposed by Alan W. Gentry and Hooker in 1988, is invalid due to the polyphyly of the lineages in the phylogenetic analysis. However, the Xiphodontidae was still found to compose part of a wider clade with the three other groups.[24] He also proposed that Leptotheridium, previously relocated from the "Dacrytheriidae" to the Xiphodontidae, composes part of a paraphyletic anoplotheriid clade with the dacrytheriines Catodontherium and Dacrytherium.[27][16][24]
Description
Skull

Dacrytherium has fairly complete skull material in the case of D. ovinum since 1876 and is best known for its large-sized lacriminal fossa in front of its eye, or "tear pit", hence the derivation of the genus name.[5][15][28] Such a depression occurs in several extant artiodactyls and is known as the "preorbital fossa", where the preorbital gland is located in living animals.[29] The preorbital fossae, which occupy the maxillae, were suggested by Nelly Delmont in 1941 to have no close analogues even amongst other mammals with preorbital fossae.[30] The infraorbital foramen are close to the orbits of the eyes and are connected to the preorbital fossa as a sac-like extension.[15] The deep preorbital fossa is present in all species except D. elegans, whose preorbital fossa has a different morphology.[31]
Jerry J. Hooker suggested that the trait supports the possibility of the D. elegans-D. saturnini and D. priscum-D. ovinum lineages having independently acquired the trait.[32] The deep preorbital fossa, well-pronounced in the genus, is not present in the anoplotheriines Anoplotherium and Diplobune. Historically the trait, along with the fusion of the front internal cusps in the lower molar teeth, were used to justify Dacrytherium as being a different evolutionary lineage from other anoplotheres. Otherwise, however, the skull of Dacrytherium is so similar to those of the anoplotheriines that Helga Sharpe Pearson questioned Dacrytherium being considered "evolutionarily separate" from anoplotheriines in 1927.[33][34]
The modern-day diagnoses of the dacrytheriine include a low-positioned roof of the cranium, an elongated and transversely developed muzzle with a wide and empty preorbital space, a lack of any postorbital bar structure, and the mastoid part of the temporal bone being exposed in the outer area between the exoccipital part of the occipital bone and the squamosal bone.[16] The skull of Dacrytherium is overall similar in structure to Anoplotherium with one main difference being the robust build of the larger skull of Anoplotherium for supporting stronger muscles. Pearson stressed that such differences do not separate the close affinities of the two genera, similar to how larger species of Mixtotherium are still close in affinity to smaller species despite their different builds or how tiger skulls are still similar to those of house cats.[34]
The skull of D. ovinum is triangular in shape and elongated in terms of the front area, while the back area is fusiform, meaning that it has a spindle-like shape with a wide middle area. It has a strong and high sagittal crest that is formed by the suture of the two parietal bones. The crest joins both the external occipital protuberance and upper nuchal lines at right angles. The zygomatic arch is thin and very prominent that it can stand out amongst the skull. The supraoccipital (or upper part of the occipital) has a shortened width but extends through the posterior edge of the skull until the end of the paroccipital process (or projection). The parietal bones are united by curvy (or sinuous) sutures to the supraoccipital, frontal, temporal, and alisphenoid bones. The sagittal crest stands at their midline and extends at the front area of the skull. The crest is high in correlation with both wide temporal fossae connected with the zygomatic bone and a narrow occiput with well-developed outer occipital crests that descend to the paroccipital processes. The frontal bones are at first narrow but quickly widen, are pierced by supraorbital foramen, and laterally border the orbits.[30][34]
The dacrytheriine has a broad mastoid part of the temporal bone for separating the exoccipital and squamosal bones that begins as a narrow bone strip on the occiput bone's edge, equivalent in position with the squamosal suture. The mastoid then broadens on the underside, down to the outer paroccipital process, as a sharp ridge then divides it into an upper portion and lower portion. The ridge is behind the mastoid's suture with a small process of the squamosal bone after the eardrum. The tympanohyal pit of the hyoid arch is in front of the stylomastoid foramen and behind the mastoid. The pit is positioned in an angle between the tympanic (eardrum) neck and the rear area of the tympanic part of the temporal bone.[34]
In the mandible, the mandibular condyle is high and above the edge of the alveolar process. The coronoid process of the mandible is separated from the articulating condyle. The external face of the mandible is hollowed from a deep masseteric fossa, and under the condyle is a clear pterygoid processes of the sphenoid. On the internal face, the angle of the mandible contains two facet joints for where the medial pterygoid muscle ends. The mandibular fossa is large, shallow, and limited in its back area by a pronounced process from the postglenoid process of the squamosal.[30] The postglenoid process is narrow and pierced from behind by a large postglenoid foramen, one of the main skull passageways for veins.[34]
Endocast anatomy

Based on two brain endocasts of the genus, one of D. ovinum and another of D. cf. ovinum, Colette Decheaux determined that their brains lacked flexure parts and had a lowered shape. Many of the brain features of the genus were identified by the palaeoneurologist as being typical of anoplotheriids. The rhinal sulcus is not in any angled shape, and the neocortex's furrows do not form circle arcs but instead form "longitudinal" arcs. The foremen of he lower face of the brain are arranged distant from each other and in order.[35]
The cerebellum of the brain is well-developed in terms of the cerebral hemispheres, is separated by a deep and large depression, and has a protruding cerebellar vermis with a well-pronounced primary fissure of cerebellum for emphasizing a greatly developed paleocerebellum. Similarly, the neocerebellum is large but longer than the paleocerebellum and is curvy because it tilts slightly to the right, then returns back, and finally reverses back to the sagittal plane of the brain. The transverse swellings in relief are marked on the neocerebellum. The cerebellar hemispheres are lower in position than the vermis and contains a large superior petrosal sinus on the surface in an arclike form, which is attached to the base of the neocerebellum but gradually moves away from it in the front area. It is above an extensive, irregular, and convex surface which, at the front area, detaches from the flocculus of the cerebellum.[35]
In the cerebrum, its cerebral hemispheres quickly enlarge from a quarter of their lengths at their front end then gradually reach their maximum widths at about four-fifths of their lengths. To their fronts is a narrow and cut short frontal lobe as well as a long and detached temporal lobe at their upper surfaces. The rhinal fissure on the cerebrum, located on the upper face of the brain based on the cast, sets the boundaries of a small neocortex of a large rhinencephalon that is not clearly visible in upper views. The fissure is halted on the left side and continues on the right, but it has a distinctive trace of its two portions, known as the anterior rhinal and the posterior rhinal. The anterior rhinal is marked deep and is strongly convex on is underside, with the neocortex projecting above it and overhanging the rhinecephalon so that only a small amount of the neocortex is visible. The posterior rhinal, although also well-marked, is not as deep as its counterpart, is rectilinear in shape of its front portion, and is raised in its rear portion towards its upper surface. The rhinencephalon has a large piriform cortex in both length and height when observed laterally.[35]
The neocortex is smoothened, although the texture is not necessarily identical on the left and right. A short and rectilinear furrow, distant from the sagittal sinus, runs diagonally from the front area to the back area towards the medial axis of the brain. Another furrow known as the suprasylvian sulcus (or suprasylvia) is slightly convex in the lower face of the brain and extends to the right up side to an upper depression. The suprasylvia then curves and connects to a short furrow located on the frontal lobe, interpreted by Colette Dechaseaux as the coronal sulcus.[35] The elongated lateral sulcus, suprasylvia, and coronal sulcus as well as a small and oblique sulcus are said to be very similar to those of cainotherioids from their "cainotherioid plan" features.[36]
Dentition

The dental formula of Dacrytherium and other anoplotheriids is 3.1.4.33.1.4.3 for a total of 44 teeth, consistent with the primitive dental formula for early-middle Paleogene placental mammals.[37][38] Anoplotheriids have selenodont or bunoselenodont premolars and molars made for folivorous/browsing diets, consistent with environment trends in the late Eocene of Europe. The canines of the Anoplotheriidae are premolariform in shape, meaning that the canines are overall undifferentiated from other teeth like incisors. The lower premolars of the family are piercing and elongated. The upper molars are bunoselenodont in form while the lower molars have selenodont labial cuspids and bunodont lingual cuspids. The subfamily Dacrytheriinae differs from the Anoplotheriinae based on the presence of a third cusp between the metaconid and entoconid of the lower molars and having molariform premolars with crescent-shaped paraconules.[18]
Dacrytherium has various specific dental diagnoses, some of which are similar to other anoplotheriids and some others of which are unique. Its upper incisors (I1-I3) are triangular in shape. The canines (C) are undifferentiated, typical of the Anoplotheriidae. The P1-P3 are elongated in size and have poorly-developed lingual lobes (or divisions). The P4 is also triangular and has a crescent-shaped lingual cusp. The P1-P3 are narrow and sharp while the P4 has a metaconid cusp that is distolingual in position to the protoconid cusp and has a weak paraconid cusp that is divided into two branches. The molars of Dacrytherium are "pentacuspidate", meaning that they have five cusps. In them, the parastyle cusp is connected to a prominent parastyle cusp, the labial sides of the paracone cusp and metacone cusp slightly ridged, and mesostyle cusps are loop-shaped. The lower molars each have two labial, cresent-shaped cusps and three lingual cusps for a total of five, with the postcristid and paracristid cusps extending lingually.[16]
In regards to the occlusion of teeth, the cusps of the lower teeth fit easily into the depressions of the upper teeth, a trait apparently well-pronounced especially in its premolars. The occlusion of Dacrytherium, according to Delmont, is similar to those of the Suidae, the main difference being that Dacrytherium lacks cutting-edge teeth.[30]
Limbs

Due to the lack of clear evidence of the phalanges of Dacrytherium as opposed to its relatives Anoplotherium and Diplobune, speculations of the number of fingers it had ranged from three fingers[39][40] to four fingers[33][41] in the 19th-20th centuries, with no clear modern evaluations of its postcranial evidence in terms of its limbs. While not rich in postcranial evidence like Anoplotherium or Diplobune, Dacrytherium is known from some limb bones, including astragali in the cases of D. elegans and D. ovinum.[25][24]
Several of such fossil materials were first described by Depéret in 1917. The calcaneum, he pointed out, was short and compressed across its back tuberosity which leads to a sharp and thick slant with a rounded head. He also attributed an astragalus to the genus and described it as being narrow plus slender in shape similar to those of the anthracothere Elomeryx and the wild boar (Sus scrofa).[4] However, in 1947, Jean Viret and J. Prudant stated that the astragalus actually belonged not to Dacrytherium but Choeropotamus while that previously attributed to the latter was reclassified as belonging to the former.[42]
The astragalus previously attributed to Choeropotamus was described by Depéret as having a twisted appearance vertically, which he said distinguishes it from the rectilinear-shaped astragali of ruminants. The bone is short and stocky in proportions, making its appearance very similar to that of Anthracotherium, the main differences setting it from the astragalus of Anthracotherium being the compressed area of the back joint and the slanting of the ridge dividing the two articular surfaces for the scaphoid and thigh.[4] Viret and Prudant considered that the astragalus reclassified to Dacrytherium had a large width, unequal lips of the tibial trochlea, a broad sustentacular facet joint, a digital pulley limited to the front area, and a deep cavity on the bone's external face. These traits, they determined, were typical of the Anoplotheriidae, leading them to favor Dacrytherium belonging to the family. Despite the size of the astragalus being large (specifically larger than those of Choeropotamus), they felt that it belonged to Dacrytherium.[42]
The morphology of the astragalus of Dacrytherium being similar to the astragali of the anoplotheriines Anoplotherium and Diplobune, as originally proposed by Viret and Prudant, was supported by Jean-Noël Martinez and Jean Sudre in 1995. They reported that the astragalus was proportionally wide and stocky and that the sustentacular facet is extensive compared to the dichobunid Messelobunodon and the suoid Doliochoerus. In contrast to the concave facet of Anoplotherium and Diplobune, that of Dacrytherium is flat to slightly convex. Because of their unique morphologies, Anoplotherium, Diplobune, and Dacrytherium had many characteristics with no modern analogues.[43]
Viret and Prudant also observed an incomplete radius that is well-preserved on the proximal end, which they said had three parts: the middle area that is hollowed out for articulation with the capitulum of the humerus and two other outer areas that form planes inclined in opposite directions. Its condylar facet is more developed compared to the trochlear facet and slopes both outwards and forward. These traits are consistent with the anatomical structures of typical anoplotheriids, corresponding to a level of mobility of the forelimb unusual for artiodactyls. They determined that one of the two radii belonged to D. ovinum and not Leptotheridium because the latter genus is smaller than the former species. The second proximal end of a radius, which they attributed to Catodontherium, differs from that of Dacrytherium by a transversely enlarged appearance, a wider end area, and a more primitive form in how less differentiated it is compared to Dacrytherium.[42]
Size
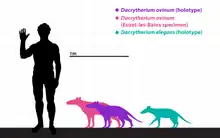
Since 1917, palaeontologists like Depéret in 1917 noticed size differences in species of Dacrytherium based on tooth sizes. Depéret explained that D. saturnini was small-sized similar to D. elegans and differed from D. ovinum only based on the smaller dimensions of its molars. D. saturnini and D. ovinum were parallel to each other in ranges and likely represented different branches of the genus, both then going extinct without leaving any descendants.[42] Depéret's argument was extended further by Sudre in 1978, who stated that D. saturnini and D. ovinum were the largest species as well as the latest of their lineages, therefore composing of the D. elegans-D. saturnini and D. priscum-D. ovinum lineages.[44][16] In 1988, however, Sudre changed his mind and determined that because D. elegans reached maximum molarization, it could not have been the ancestor of D. saturnini, potentially leaving the descendant of the latter unknown.[45][31]
Martinez and Sudre followed up with weight estimates for D. saturnini amongst other Paleogene artiodactyls in 1995 based on the dimensions of their astragali and M1 teeth. The astragali are common bones in fossil assemblages due to their reduced vulnerability to fragmentation as a result of their stocky shape and compact structure, explaining their choice for using it. The two weight estimates of D. saturnini from the locality of Sainte Néboule (MP18) yielded different results, the M1 giving the body mass of 7.51 kg (16.6 lb) and the astragalus yielding 13.187 kg (29.07 lb). The researchers considered that the body weight of D. saturnini from the M1 is an underestimate compared to the result from the astragalus.[43]
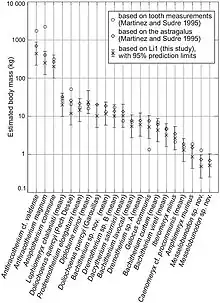
In 2014, Takehisa Tsubamoto reexamined the relationship between astragalus size and estimated body mass based on extensive studies of extant terrestrial mammals, reapplying the methods to Paleogene artiodactyls previously tested by Sudre and Martinez. The researcher used linear measurements and their products with adjusted correction factors. The recalculations resulted in somewhat lower estimates compared to the 1995 results (with the exception of Diplobune minor, which as a shorter astragalus proportion than most other artiodactyls), displayed in the below graph:[46]
Palaeobiology

The palaeobiologies of anoplotheriids including the dacrytheriine Dacrytherium are uncertain. While palaeontologists had historically established anoplotheriids as having unusual postcranial morphologies with no modern analogues amongst artiodactyls, their behaviours are still unknown as postcranial evidence for most anoplotheriids, including Dacrytherium, remain scarce.[42][43] Modern-day hypotheses range from arborealism in the case of Diplobune[47][48] to bipedalism in the case of Anoplotherium (Jerry J. Hooker also speculated the possibility of other anoplotheriids sharing similar behaviours).[25] Alternatively, Hooker in 1986 suggested that Dacrytherium may have been a purely ground-dwelling folivore.[49]
Hooker in 1986 also pointed out that Dacrytherium and Mixtotherium, despite belonging to different artiodactyl families, had similar dentitions based on their low-crowned and strongly selenodont molars.[49] The Dacrytheriinae and Anoplotheriinae are thought to have belonged to the selenodont dentition group of endemic European Paleogene artiodactyls, meaning that they were likely folivorous browsers.[50]
Palaeoecology
Middle Eocene

For much of the Eocene, a hothouse climate with humid, tropical environments with consistently high precipitations prevailed. Modern mammalian orders including the Perissodactyla, Artiodactyla, and Primates (or the suborder Euprimates) appeared already by the early Eocene, diversifying rapidly and developing dentitions specialized for folivory. The omnivorous forms mostly either switched to folivorous diets or went extinct by the middle Eocene (47–37 Ma) along with the archaic "condylarths". By the late Eocene (approx. 37–33 Ma), most of the ungulate form dentitions shifted from bunodont cusps to cutting ridges (i.e. lophs) for folivorous diets.[51][52]
Land-based connections to the north of the developing Atlantic Ocean were interrupted around 53 Ma, meaning that North America and Greenland were no longer well-connected to western Europe. From the early Eocene up until the Grande Coupure extinction event (56 Ma - 33.9 Ma), the western Eurasian continent was separated into three landmasses, the former two of which were isolated by seaways: western Europe (an archipelago), Balkanatolia, and eastern Eurasia (Balkanatolia was in between the Paratethys Sea of the north and the Neotethys Ocean of the south).[17] The Holarctic mammalian faunas of western Europe were therefore mostly isolated from other continents including Greenland, Africa, and eastern Eurasia, allowing for endemism to occur within western Europe.[52] The European mammals of the late Eocene (MP17 - MP20) were mostly descendants of endemic middle Eocene groups as a result.[53]

The first undisputed appearance of Dacrytherium was by MP13 in the form of the species D. cf. elegans.[54] D. priscum, meanwhile, is known exclusively from the Swiss deposit of Egerkingen, which dates back to MP14.[31] By MP13, the dacrytheriine coexisted with perissodactyls (Palaeotheriidae, Lophiodontidae, and Tapiroidea), non-endemic artiodactyls (Dichobunidae and Tapirulidae), endemic European artiodactyls (Choeropotamidae (possibly polyphyletic, however), Cebochoeridae, Mixtotheriidae, and other members of Anoplotheriidae), and primates (Adapidae). Both the Amphimerycidae and Xiphodontidae made their appearances by the level MP14.[50][22][55] The stratigraphic ranges of the early species of Dacrytherium also overlapped with marsupials (Herpetotheriidae), cimolestans (Pantolestidae, Paroxyclaenidae), rodents (Ischyromyidae, Theridomyoidea, Gliridae), eulipotyphlans, bats, apatotherians, carnivoraformes (Miacidae), and hyaenodonts (Hyainailourinae, Proviverrinae).[56] Other MP13-MP14 sites have also yielded fossils of turtles and crocodylomorphs,[57] and MP13 sites are stratigraphically the latest to have yielded remains of the Gastornithidae and Palaeognathae.[58]
In the level MP13, D. cf. elegans fossils cooccurred with those of many other mammals such as the herpetotheriid Amphiperatherium, miacid Quercygale, proviverrine Proviverra, equoid Hallensia, palaeotheres Propalaeotherium and Plagiolophus, lophiodont Lophiodon, choeropotamids (Haplobunodon, Rhagatherium and Amphirhagatherium), and the cebochoerid Cebochoerus.[54][56][16]
Undisputed fossil remains of D. elegans occur in several sites of France and Switzerland that date back to MP16, such as Mormont Eclépens, Le Bretou, and Robiac.[31] The locality of Robiac indicates that D. elegans coexisted with similar mammal faunas as earlier species of the genus, such as the herpetotheriids Peratherium and Amphiperatherium, hyaenodonts Paroxyaena and Cynohyaenodon, miacids Paramiacis and Quercygale, palaeotheres (Palaeotherium, Plagiolophus, Anchilophus), lophiodont Lophiodon, cebochoerids Cebochoerus and Acotherulum, choeropotamid Choeropotamus, dichobunid Mouillacitherium, robiacinid Robiacina, xiphodonts (Xiphodon, Dichodon, and Haplomeryx), amphimerycid Amphimeryx, and other anoplotheriids Catodontherium and Robiatherium.[56] Fossil localities such as the Creechbarrow Limestone formation in England, an MP16 locality where some D. elegans fossils were uncovered, suggest a subtropical climate that could support closed forested environments for arboreal animals and animals with folivorous and/or frugivorous diets.[49][59]
By MP16, a faunal turnover occurred, marking the disappearances of the lophiodonts and European tapiroids as well as the extinctions of all European crocodylomorphs except for the alligatoroid Diplocynodon.[22][57][60][61] The causes of the faunal turnover have been attributed to a shift from humid and highly tropical environments to drier and more temperate forests with open areas and more abrasive vegetation. The surviving herbivorous faunas shifted their dentitions and dietary strategies accordingly to adapt to abrasive and seasonal vegetation.[62][63] The environments were still subhumid and full of subtropical evergreen forests, however. The Palaeotheriidae was the sole remaining European perissodactyl group, and frugivorous-folivorous or purely folivorous artiodactyls became the dominant group in western Europe.[64][50]
Late Eocene
In the late Eocene, there were two species of Dacrytherium: D. ovinum and D. saturnini. D. ovinum ranges stratigraphically from MP17a to MP18 while D. saturnini ranges from MP18 to MP19.[31][54] Both lineages largely coexisted with the same artiodactyl families as well as the Palaeotheriidae within western Europe,[56] although the Cainotheriidae and the derived anoplotheriids Anoplotherium and Diplobune all made their first fossil record appearances by MP18.[16][65] In addition, several migrant mammal groups had reached western Europe by MP17a-MP18, namely the Anthracotheriidae, Hyaenodontinae, and Amphicyonidae.[56] In addition to snakes, frogs, and salamandrids, rich assemblage of lizards are known in western Europe as well from MP16-MP20, representing the Iguanidae, Lacertidae, Gekkonidae, Agamidae, Scincidae, Helodermatidae, and Varanoidea, most of which were able to thrive in the warm temperatures of western Europe.[66]
The MP18 locality of La Débruge of France indicates that D. saturnini coexisted with a wide variety of mammals, namely the herpetotheriid Peratherium, rodents (Blainvillimys, Theridomys, Plesiarctomys, Glamys), hyaenodonts (Hyaenodon and Pterodon), amphicyonid Cynodictis, palaeotheres (Plagiolophus, Anchilophus, Palaeotherium), dichobunid Dichobune, choeropotamid Choeropotamus, cebochoerids Cebochoerus and Acotherulum, anoplotheriids Anoplotherium and Diplobune, tapirulid Tapirulus, xiphodonts Xiphodon and Dichodon, cainothere Oxacron, amphimerycid Amphimeryx, and anthracothere Elomeryx.[56]
See also
References
- ↑ Filhol, Henri (1876). "Mammifères fossiles nouveaux provenant des dépôts de phosphate de chaux du Quercy". Comptes Rendus de l'Académie des Sciences. 82: 288–289.
- ↑ Gervais, Paul (1876). "Mammifères appartenant à l'ordre des Bisulques.". Zoologie et paléontologie générales 2. série Nouvelles recherches sur les animaux vertébrés dont on trouve les ossements enfouis dans le sol et sur leur comparaison avec les espèces actuellement existantes. Arthus Bertrand. pp. 42–63.
- 1 2 Palmer, Theodore Sherman (1904). "A List of the Genera and Families of Mammals". North American Fauna (23). doi:10.3996/nafa.23.0001.
- 1 2 3 Depéret, Charles (1917). Monographie de la faune de mammifères fossiles du Ludien inférieur d'Euzet-les-Bains (Gard). Lyon A. Rey.
- 1 2 3 Filhol, Henri (1877). "Recherches sur les phosphorites du Quercy: etude des fossiles qu'on y rencontre et spécialement des mammifères". Annales des sciences géologiques. 8: 217–225.
- ↑ Filhol, Henri (1884). "Pachydermes". Descriptions de Quelques Mammifères Fossiles des Phosphorites du Quercy. Vialelle Printing Company and Co. pp. 33–34.
- ↑ Filhol, Henri (1880). "Sur la dècouverte de Mammifères nouveaux dans les dèpôts de phosphate de chaux du Quercy". Comptes rendus hebdomadaires des séances de l'Académie des sciences. 90: 1579–1580.
- ↑ Jentink, Fredericus Anna (1880). "6. Mammalia". Zoologischer Jahresbericht. 4: 259.
- ↑ Owen, Richard (1857). "Description of the Lower Jaw and Teeth of an Anoplotherioid quadruped (Dichobune ovina, Ow.) of the size of the Xiphodon gracilis, Cuv., from the Upper Eocene Marl, Isle of Wight". The Quarterly Journal of the Geological Society of London. 13 (1–2): 254–260. doi:10.1144/GSL.JGS.1857.013.01-02.38. S2CID 130007945.
- ↑ Lydekker, Richard (1885). Catalogue of the fossil Mammalia in the British museum, (Natural History): Part II. Containing the Order Ungulata, Suborder Artiodactyla. Order of the Trustees, London.
- ↑ von Zittel, Karl Alfred (1891–1893). Handbuch der Palaeontologie. I. Abtheilung. Palaeozoologie von Karl A. Zittel. IV. Band. (Mammalia). R. Oldenbourg. pp. 370–374.
- ↑ Lydekker, Richard (1892). "On Dacrytherium ovinum from the Isle of Wight and Quercy (Read November 11, 1891)". The Quarterly Journal of the Geological Society of London. 48: 1–4. doi:10.1144/GSL.JGS.1892.048.01-04.01. S2CID 219222697.
- ↑ Stehlin, Hans Georg (1908). "Die Säugetiere des schweizerischen Eocaens. Sechster Teil: Choeropotamus – Cebochoerus – Choeromorus – Haplobunodon – Rhagatherium – Mixtotherium". Abhandlungen der Schweizerischen Paläontologischen Gesellschaft. 35.
- ↑ Rütimeyer, Ludwig (1891). "II. Ungulata Paridigitata". Abhandlungen der Schweizerischen paläontologischen Gesellschaft. 18: 61–62.
- 1 2 3 Stehlin, Hans Georg (1910). "Die Säugertiere des schweizerischen Eocaens. Sechster Teil: Catodontherium – Dacrytherium – Leptotherium – Anoplotherium – Diplobune – Xiphodon – Pseudamphimeryx – Amphimeryx – Dichodon – Haplomeryx – Tapirulus – Gelocus. Nachträge, Artiodactyla incertae sedis, Schlussbetrachtungen über die Artiodactylen, Nachträge zu den Perissodactylen". Abhandlungen der Schweizerischen Paläontologischen Gesellschaft. 36.
- 1 2 3 4 5 6 7 8 9 Erfurt, Jörg; Métais, Grégoire (2007). "Endemic European Paleogene Artiodactyls". In Prothero, Donald R.; Foss, Scott E. (eds.). The Evolution of Artiodactyls. Johns Hopkins University Press. pp. 59–84.
- 1 2 Licht, Alexis; Métais, Grégoire; Coster, Pauline; İbilioğlu, Deniz; Ocakoğlu, Faruk; Westerweel, Jan; Mueller, Megan; Campbell, Clay; Mattingly, Spencer; Wood, Melissa C.; Beard, K. Christopher (2022). "Balkanatolia: The insular mammalian biogeographic province that partly paved the way to the Grande Coupure". Earth-Science Reviews. 226: 103929. Bibcode:2022ESRv..22603929L. doi:10.1016/j.earscirev.2022.103929.
- 1 2 Badiola, Ainara; De Vicuña, Nahia Jiménez; Perales-Gogenola, Leire; Gómez-Olivencia, Asier (2023). "First clear evidence of Anoplotherium (Mammalia, Artiodactyla) in the Iberian Peninsula: an update on the Iberian anoplotheriines". The Anatomical Record: Advances in Integrative Anatomy and Evolutionary Biology. doi:10.1002/ar.25238. PMID 37221992. S2CID 258864256.
- 1 2 Weppe, Romain; Blondel, Cécile; Vianey-Liaud, Monique; Pélissié, Thierry; Orliac, Maëva Judith (2020). "A new Cainotherioidea (Mammalia, Artiodactyla) from Palembert (Quercy, SW France): Phylogenetic relationships and evolutionary history of the dental pattern of Cainotheriidae". Palaeontologia Electronica (23(3):a54). doi:10.26879/1081. S2CID 229490410.
- 1 2 Luccisano, Vincent; Sudre, Jean; Lihoreau, Fabrice (2020). "Revision of the Eocene artiodactyls (Mammalia, Placentalia) from Aumelas and Saint-Martin-de-Londres (Montpellier limestones, Hérault, France) questions the early European artiodactyl radiation". Journal of Systematic Palaeontology. 18 (19): 1631–1656. Bibcode:2020JSPal..18.1631L. doi:10.1080/14772019.2020.1799253. S2CID 221468663.
- ↑ Sudre, Jean; Lecomte, Gérard (2000). "Relations et position systématique du genre Cuisitherium Sudre et al., 1983, le plus dérivé des artiodactyles de l'Éocène inférieur d'Europe" (PDF). Geodiversitas. 22 (3): 415–432.
- 1 2 3 Franzen, Jens Lorenz (2003). "Mammalian faunal turnover in the Eocene of central Europe". Geological Society of America Special Papers. 369: 455–461. doi:10.1130/0-8137-2369-8.455. ISBN 9780813723693.
- 1 2 Cuesta, Miguel-Ángel; Badiola, Ainara (2009). "Duerotherium sudrei gen. et sp. nov., a New Anoplotheriine Artiodactyl from the Middle Eocene of the Iberian Peninsula". Journal of Vertebrate Paleontology. 29 (1): 303–308. Bibcode:2009JVPal..29..303C. doi:10.1671/039.029.0110. JSTOR 20491092. S2CID 55546022.
- 1 2 3 4 5 Weppe, Romain (2022). Déclin des artiodactyles endémiques européens, autopsie d'une extinction (Thesis) (in French). University of Montpellier.
- 1 2 3 Hooker, Jerry J. (2007). "Bipedal browsing adaptations of the unusual Late Eocene–earliest Oligocene tylopod Anoplotherium (Artiodactyla, Mammalia)". Zoological Journal of the Linnean Society. 151 (3): 609–659. doi:10.1111/j.1096-3642.2007.00352.x.
- 1 2 Weppe, Romain; Blondel, Cécile; Vianey-Liaud, Monique; Escarguel, Gilles; Pélissié, Thierry; Antoine, Pierre-Olivier; Orliac, Maëva Judith (2020). "Cainotheriidae (Mammalia, Artiodactyla) from Dams (Quercy, SW France): phylogenetic relationships and evolution around the Eocene–Oligocene transition (MP19–MP21)" (PDF). Journal of Systematic Palaeontology. 18 (7): 541–572. Bibcode:2020JSPal..18..541W. doi:10.1080/14772019.2019.1645754. S2CID 202026238.
- ↑ Ruiz-Colmenares, Miguel Ángel Cuesta (1998). "Presencia de Leptotheridium (Dacrytheriidae, Artiodactyla, Mammalia) en el yacimiento eocénico de Caenes (Cuenca del Duero, Salamanca, España)". Studia Geologica Salmanticensia. 34: 69–78.
- ↑ Lydekker, Richard (1894). "The Ancient Mammals of Britain. III.- The Lower Tertiary Period.". Knowledge: A Monthly Record of Science. Vol. 17. Knowledge Publishing Company. pp. 221–223.
- ↑ Rehorek, Susan J.; Hillenius, Willem J.; Kennaugh, John; Chapman, Norma (2005). The gland and the sac — the preorbital apparatus of muntjacs. Chemical Signals in Vertebrates 10. pp. 152–158. doi:10.1007/0-387-25160-X_19.
- 1 2 3 4 Delmont, Nelly (1941). "Un Mammifère Artiodactyle de l'Eocène: le Dacrytherium". Annales de Paléontologie. 29: 29–50.
- 1 2 3 4 5 Ruiz-Colmenares, Cuesta; Ángel, Miguel; Soler, Lluís Checa; Casanovas-Cladellas, María Lourdes (2006). "Artiodáctilos del yacimiento de Sossís (Eoceno superior, Cuenca Prepirenaica, Península Ibérica)". Revista Española de Paleontología. 21 (2): 123–144. doi:10.7203/sjp.21.2.20486. hdl:10366/82129.
- ↑ Hooker, Jerry J. (2000). "Eocene Mammal Faunas of Mormont, Switzerland: Systematic Revision and Resolution of Dating Problems". Kommission der Schweizerischen Paläontologischen Abhandlungen. 120: 92–94.
- 1 2 Earle, Charles (1896). "Notes on the Fossil Mammalia of Europe, V – The Phylogeny of Anoplotherium". The American Naturalist. 30: 665–668.
- 1 2 3 4 5 Pearson, Helga Sharpe (1927). "On the Skulls of Early Tertiary Suidae, together with an Account of the Otic Region in Some Other Primitive Artiodactyla". Philosophical Transactions of the Royal Society of London. Series B, Containing Papers of a Biological Character. 215 (421–430): 440–445. doi:10.1098/rstb.1927.0009.
- 1 2 3 4 Decheaux, Colette (1969). "Moulages endocrâniens d'artiodactyles primitifs. Essai sur l'histoire du néopallium". Annales de Paléontologie. 55: 195–248.
- ↑ Orliac, Maeva J.; Maugoust, Jacob; Balcarcel, Ana; Gilissen, Emmanuel (2023). "Paleoneurology of Artiodactyla, an Overview of the Evolution of the Artiodactyl Brain" (PDF). In Dozo; Paulina-Carabajal, Ariana; Macrini, Thomas E.; Walsh, Stig (eds.). Paleoneurology of Amniotes. Springer Cham. pp. 507–555. doi:10.1007/978-3-031-13983-3_13. ISBN 978-3-031-13982-6.
- ↑ von Zittel, Karl Alfred (1925). Schlosser, Max (ed.). Text-Book of Paleontology. Volume III. Mammalia. Macmillan and Co. Limited. pp. 179–180.
- ↑ Lihoreau, Fabrice; Boisserie, Jean-Renaud; Viriot, Laurent; Brunet, Michel (2006). "Anthracothere dental anatomy reveals a late Miocene Chado-Libyan bioprovince". Proceedings of the National Academy of Sciences. 103 (23): 8763–8767. Bibcode:2006PNAS..103.8763L. doi:10.1073/pnas.0603126103. PMC 1482652. PMID 16723392.
- ↑ Friant, M. (1967). "La morphologie des molaires chez les Ruminants (Ongulés artiodactyles sélénodontes) d'Europe. Son évolution phylogénique". Acta Zoologica. 48 (1–2): 87–101. doi:10.1111/j.1463-6395.1967.tb00133.x.
- ↑ Martinez, Jean-Noël (1991). L'astragale chez quelques artiodactyles du Paléogène: morphologie comparée, morphométrie, aspect fonctionnel, intérêt systématique (Thesis) (in French). University of Montpellier.
- ↑ Boule, Marcellin; Piveteau, Jean (1935). "Une patte antérieure de Diplobune". Archives du Muséum national d'histoire naturelle. 6. 12: 253–258.
- 1 2 3 4 5 Viret, Jean; Prudant, J. (1947). "Observations sur quelques caracteres anatomiques des Dacrytheridés". Extraits des Comptes Rendus de la Société Géologique de France: 26–27.
- 1 2 3 Sudre, Jean; Martinez, Jean-Noël (1995). "The astragalus of Paleogene artiodactyls: comparative morphology, variability and prediction of body mass". Lethaia. 28 (3): 197–209. Bibcode:1995Letha..28..197M. doi:10.1111/j.1502-3931.1995.tb01423.x.
- ↑ Sudre, Jean (1978). Les Artiodactyles de l'Eocéne moyen et supérieur d'Europe occidentale. University of Montpellier.
- ↑ Sudre, Jean (1988). "Le gisement du Bretou (Phosphorites du Quercy, Tarn-et-Garonne, France) et sa faune des vertebres de l'Eocene superieur: 7. Artiodactyles". Palaeontographica. Abteilung A, Paläozoologie, Stratigraphie. 205 (1–6): 129–154.
- ↑ Tsubamoto, Takehisa (2014). "Estimating body mass from the astragalus in mammals". Acta Palaeontologica Polonica. 59 (2): 259–265. doi:10.4202/app.2011.0067. S2CID 54686160.
- ↑ Sudre, Jean (1982). "Interprétation de la denture et description des éléments du squelette appendiculaire de l'espèce Diplobune minor (Filhol 1877); apports à la connaissance de l'anatomie des Anoplotheriinae Bonaparte 1850". In Mazin, J.M.; Salmon, E. (eds.). Actes du Symposium paléontologique Georges Cuvier, Montbéliard - France, 1982: communications données à l'occasion du cent cinquantième anniversaire de la mort de Georges Cuvier, du 25 octobre au 28 octobre 1982, au Musée du Château. Le Musée du Château. pp. 439–458.
- ↑ Métais, Grégoire (2014). On the "thumb" of anoplotheriins: a 3D comparative study of the hand of Anoplotherium and Diplobune. Swiss Geoscience Meeting 2014.
- 1 2 3 Hooker, Jerry J. (1986). "Mammals from the Bartonian (middle late Eocene) of the Hampshire Basin, southern England". Bulletin of the British Museum (Natural History) Geology. 39 (4): 191–478.
- 1 2 3 Blondel, Cécile (2001). "The Eocene-Oligocene ungulates from Western Europe and their environment" (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 168 (1–2): 125–139. Bibcode:2001PPP...168..125B. doi:10.1016/S0031-0182(00)00252-2.
- ↑ Eronen, Jussi T.; Janis, Christine M.; Chamberlain, Charles Page; Mulch, Andreas (2015). "Mountain uplift explains differences in Palaeogene patterns of mammalian evolution and extinction between North America and Europe". Proceedings of the Royal Society B: Biological Sciences. 282 (1809): 20150136. doi:10.1098/rspb.2015.0136. PMC 4590438. PMID 26041349.
- 1 2 Maitre, Elodie (2014). "Western European middle Eocene to early Oligocene Chiroptera: systematics, phylogeny and palaeoecology based on new material from the Quercy (France)". Swiss Journal of Palaeontology. 133 (2): 141–242. Bibcode:2014SwJP..133..141M. doi:10.1007/s13358-014-0069-3. S2CID 84066785.
- ↑ Badiola, Ainara; Perales-Gogenola, Leire; Astibia, Humberto; Suberbiola, Xabier Pereda (2022). "A synthesis of Eocene equoids (Perissodactyla, Mammalia) from the Iberian Peninsula: new signs of endemism". Historical Biology. 34 (8): 1623–1631. Bibcode:2022HBio...34.1623B. doi:10.1080/08912963.2022.2060098. S2CID 248164842.
- 1 2 3 Schmidt-Kittler, Norbert; Godinot, Marc; Franzen, Jens L.; Hooker, Jeremy J. (1987). "European reference levels and correlation tables". Münchner geowissenschaftliche Abhandlungen A10. Pfeil Verlag, München. pp. 13–31.
- ↑ Bai, Bin; Wang, Yuan-Qing; Theodor, Jessica M.; Meng, Jin (2023). "Small artiodactyls with tapir-like teeth from the middle Eocene of the Erlian Basin, Inner Mongolia, China". Frontiers in Earth Science. 11: 1–20. Bibcode:2023FrEaS..1117911B. doi:10.3389/feart.2023.1117911.
- 1 2 3 4 5 6 Aguilar, Jean-Pierre; Legendre, Serge; Michaux, Jacques (1997). "Synthèses et tableaux de corrélations". Actes du Congrès Bio-chroM'97. Mémoires et Travaux de l'EPHE Institut de Montpellier 21 (in French). École Pratique des Hautes Études-Sciences de la Vie et de la Terre, Montpellier. pp. 769–850.
- 1 2 Martin, Jeremy E.; Pochat-Cottilloux, Yohan; Laurent, Yves; Perrier, Vincent; Robert, Emmanuel; Antoine, Pierre-Olivier (2022). "Anatomy and phylogeny of an exceptionally large sebecid (Crocodylomorpha) from the middle Eocene of southern France". Journal of Vertebrate Paleontology. 42 (4). Bibcode:2022JVPal..42E3828M. doi:10.1080/02724634.2023.2193828. S2CID 258361595.
- ↑ Buffetaut, Eric; Angst, Delphine (2014). Stratigraphic Distribution of Large Flightless Birds in the Palaeogene of Europe. STRATI 2013: First International Congress on Stratigraphy At the Cutting Edge of Stratigraphy. doi:10.1007/978-3-319-04364-7_190.
- ↑ Minwer-Barakat, Raef; Badiola, Ainara; Marigó, Judit; Moyà-Solà, Salvador (2013). "First record of the genus Microchoerus (Omomyidae, Primates) in the western Iberian Peninsula and its palaeobiogeographic implications". Journal of Human Evolution. 65 (3): 313–321. doi:10.1016/j.jhevol.2013.07.002. PMID 23916791.
- ↑ Martin, Jeremy E. (2015). "A sebecosuchian in a middle Eocene karst with comments on the dorsal shield in Crocodylomorpha". Acta Palaeontologica Polonica. 60 (3): 673–680. doi:10.4202/app.00072.2014. S2CID 54002673.
- ↑ Antunes, Miguel Telles (2003). "Lower Paleogene Crocodilians from Silveirinha, Portugal". Palaeovertebrata. 32: 1–26.
- ↑ Robinet, Céline; Remy, Jean Albert; Laurent, Yves; Danilo, Laure; Lihoreau, Fabrice (2015). "A new genus of Lophiodontidae (Perissodactyla, Mammalia) from the early Eocene of La Borie (Southern France) and the origin of the genus Lophiodon Cuvier, 1822". Geobios. 48 (1): 25–38. Bibcode:2015Geobi..48...25R. doi:10.1016/j.geobios.2014.11.003.
- ↑ Perales-Gogenola, Leire; Badiola, Ainara; Gómez-Olivencia, Asier; Pereda-Suberbiola, Xabier (2022). "A remarkable new paleotheriid (Mammalia) in the endemic Iberian Eocene perissodactyl fauna". Journal of Vertebrate Paleontology. 42 (4). Bibcode:2022JVPal..42E9447P. doi:10.1080/02724634.2023.2189447. S2CID 258663753.
- ↑ Solé, Floréal; Fischer, Valentin; Le Verger, Kévin; Mennecart, Bastien; Speijer, Robert P.; Peigné, Stéphane; Smith, Thierry (2022). "Evolution of European carnivorous mammal assemblages through the Paleogene". Biological Journal of the Linnean Society. 135 (4): 734–753. doi:10.1093/biolinnean/blac002.
- ↑ Weppe, Romain; Blondel, Cécile; Vianey-Liaud, Monique; Escarguel, Gilles; Pelissie, Thierry; Antoine, Pierre-Olivier; Orliac, Maeva J. (2020). "Cainotheriidae (Mammalia, Artiodactyla) from Dams (Quercy, SW France): phylogenetic relationships and evolution around the Eocene–Oligocene transition (MP19–MP21)" (PDF). Journal of Systematic Palaeontology. 18 (7): 541–572. Bibcode:2020JSPal..18..541W. doi:10.1080/14772019.2019.1645754. S2CID 202026238.
- ↑ Rage, Jean-Claude (2012). "Amphibians and squamates in the Eocene of Europe: what do they tell us?". Palaeobiodiversity and Palaeoenvironments. 92 (4): 445–457. Bibcode:2012PdPe...92..445R. doi:10.1007/s12549-012-0087-3. S2CID 128651937.