 | |
 | |
| Names | |
|---|---|
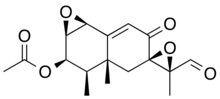
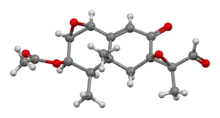
| IUPAC name
(11S)-8,12-Dioxo-1β,2β:7,11-diepoxy-7α-eremophil-9-en-3β-yl acetate | |
| Systematic IUPAC name
(1aR,2R,2′R,3R,3′S,3aR,7bS)-3′-Formyl-3,3′,3a-trimethyl-6-oxo-1a,2,3a,4,6,7b-hexahydro-3H-spiro[naphtho[1,2-b]oxirene-5,2′-oxiran]-2-yl acetate | |
| Identifiers | |
3D model (JSmol) |
|
| 3DMet | |
| ChEBI | |
| ChEMBL | |
| ChemSpider | |
| KEGG | |
PubChem CID |
|
| UNII | |
CompTox Dashboard (EPA) |
|
| |
| |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa).
Infobox references | |
Penicillin Roquefort Toxin (PR toxin) is a mycotoxin produced by the fungi Penicillium roqueforti. In 1973, PR toxin was first partially characterized by isolating moldy corn on which the fungi had grown.[1] Although its lethal dose was determined shortly after the isolation of the chemical, details of its toxic effects were not fully clarified until 1982 in a study with mice, rats, anesthetized cats and preparations of isolated rat auricles.[2]
Structure and reactivity
PR toxin contain multiple functional groups, including acetoxy (CH3COO-), aldehyde (-CHO), α,β-unsaturated ketone (-C=C-CO) and two epoxides.[3] The aldehyde group on C-12 is directly involved in the biological activity as removal leads to inactivation of the compound. The two epoxide groups do not play an important role, as removal showed no difference in activity. When exposed to air, PR toxin may decompose. How and why this happens, is however not known.[4]
Synthesis
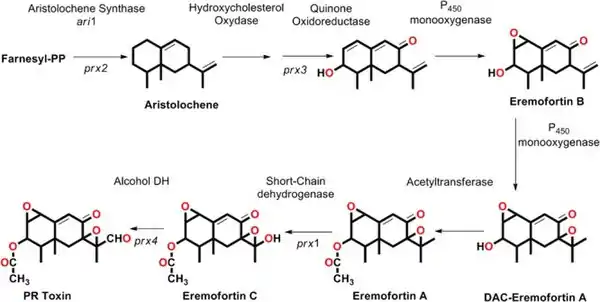
PR toxin is derived from the 15-carbon hydrocarbon aristolochene, a sesquiterpene produced from farnesyl diphosphate catalyzed by the enzyme aristolochene synthase. Aristolochene then gains an alcohol, a ketone, and an additional alkene, mediated by hydroxysterol oxidase and quinone oxidoreductase. Addition of the fused-epoxide oxygen by P450 monooxygenase gives eremofortin B. Epoxidation of the isopropenyl sidechain, again by P450 monooxygenase, and addition of the acetyl group by an acetyltransferase gives eremofortin A. A short-chain oxidoreductase oxidizes a methyl group on the side-chain to eremofortin C, the primary alcohol analog of PR toxin (incorrectly illustrated in the following diagram), which is then further oxidized by a short-chain alcohol dehydrogenase to give the aldehyde.[3]
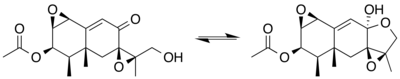
Eremofortin C has been isolated from microbial sources and found to be in a spontaneous equilibrium between an open-chain hydroxy–ketone structure and a lactol form.[5]
Metabolism
Different experiments have shown the effects of the PR toxin on liver cells in culture (in vitro) and in the liver (in vivo).[4][6][7]
In vitro
The PR toxin have caused an inhibition of incorporating amino acids, these results show that the toxin was responsible for altering the translating process. Together with some earlier experiments it has been proved that the PR toxin was indeed active on the cell metabolism.[4][6] Another interesting finding is the decreased activity of the respiratory control and oxidative phosphorylation in the (isolated) mitochondria of the liver .
Apparently the amount of polysomes wasn't the determining factor, the inhibition was not decreased by increasing the amount of polysomes. The increase of ph5 enzymes on the other hand, had a significant inhibitory effect. A higher concentration of ph5 enzymes made the inhibitory effect less effective. These findings proved that the PR toxin was not altering the polysomes but in some way disfunctions the ph5 enzymes.[6]
In vivo
When the PR toxin was directly administered to rats, the protein synthesis in the liver was not as high as it normal would be.[6] This in in vivo administration showed that the isolated cells from the rat's liver had a much lower transcriptional capacity.[4]
The process did not alter the uptake of amino acids in the liver, but exclusively the translational process was affected. The toxic effect of this toxin is as expected close with the fact that the process of protein synthesis is inhibited. However the real toxic effect could be that some required proteins aren't made in a proper amount to function in a right way.[6]
Mechanism of action
Multiple experiments have shown the different effects of PR toxin: it can cause damage to the liver and kidney, can induce carcinogenicity, and can in vivo inhibit the DNA replication, protein synthesis, and transcription.[3] Most experiments on the effect of the PR toxin focus on the inhibition of protein synthesis and impairment of the liver.[4][6][3]
The PR toxin dysfunctions the transcriptional process in the liver. The RNA polymerases I & II, the two main RNA polymerase systems in the liver, are affected by the toxin. The toxin needs no further enzymatic conversion to exert its effects on these systems.[4] The liver seems to be the most influenced organ by the PR toxin.
Toxicity
The toxicity of PR toxin was measured both intraperitoneal as well as oral. The first determined median lethal dose of pure PR toxin intraperitoneal in weanling rats was 11 mg/kg. The oral median lethal dose was 115 mg/kg.[1] The same study reported that ten minutes after an oral dose of 160 mg/kg, the animals experienced breathing problems that eventually lead to death.
Acute Rat studies (mg/kg)
- LDLo test, via oral route: 115
- LD50 test, intraperitoneal route: 11.6
- LD50 test, intravenous: 8.2
Acute Mouse studies (mg/kg)
- LD50 test, via oral route: 72
- LD50 test, intraperitoneal: 2
- LD50 test, intravenous: 2
An acute human study has yet to be done, so no LD50 test results or doses are known yet. However, there is one case report from 1982 in which toxic effects are described on a human. This person was working in a factory in which the blue cheese was produced. The mold of Penicillium roqueforti was inhaled by this person and she developed hypersensitivity pneumonitis. Because of this lung inflammation, the person experienced among other things coughing, dyspnea, reduced lung volumes and hypoxemia. Antibodies against the mold were found afterwards in serum and lavage fluid. However, the LD50 values have not been determined yet.[8]
Effects on animals
Studies of the effects on animals were done on mice, rats, anesthetized cats and preparations of isolated rat auricle. Toxic effects in mice and rats included abdominal writhing, decrease of motor activity and respiration rate, weakness of the hind leg and ataxia.
The effects were different for the different ways PR toxin was taken up. When the median lethal dose was ingested orally, the pathology was described as swollen-gas filled stomach and intestines as well as edema and congestion in the lungs. The kidney showed degenerative changes as well as hemorrhage.
If PR toxin was injected intraperitoneally, cats, mice and rats developed ascites fluid and edema of the lungs and scrotum. While intravenous injection showed, for the same animals, large volumes of pleural and pericardial volumes as well as lung edema.
In conclusion, the tissue cells and blood vessels were directly damaged by PR toxin. This caused leakage of fluid resulting among other things in edema of the lung and ascites fluid. Also, the damage on the blood vessels resulted in increased capillary permeability. This increased permeability lead to a decrease in blood volume and direct damage to the vital organs including lungs, kidneys, liver and heart.[9]
References
- 1 2 Wei, R.D.; et al. (1973-01-25). "Isolation and Partial Characterization of a Mycotoxin from Penicillium roqueforti". American Society for Microbiology. 25 (1): 111–114. doi:10.1128/am.25.1.111-114.1973. PMC 380745. PMID 4687064.
- ↑ Chen, F.C.; et al. (1982). "Acute toxicity of PR toxin, a mycotoxin from Penicillium roqueforti'". Toxicon. 20 (2): 433–441. doi:10.1016/0041-0101(82)90006-x. PMID 7080052.
- 1 2 3 4 Dubey, M.K.; et al. (2019-03-29). "PR Toxin - Biosynthesis, Genetic Regulation, Toxicological Potential, Prevention and Control Measures: Overview and Challenges". Frontiers in Pharmacology. 9: 288. doi:10.3389/fphar.2018.00288. PMC 5885497. PMID 29651243.
- 1 2 3 4 5 6 Moule, Y.; et al. (1976). "Mechanism of the inhibition of transcription by pr toxin, a mycotoxin from Penicillium roqueforti". Chemico-Biological Interactions. 14 (3–4): 207–216. doi:10.1016/0009-2797(76)90101-0. PMID 182392.
- ↑ Moreau, Serge; Cacan, Monique; Lablache-Combier, Alain (1997). "Eremofortin C, a new metabolite obtained from Penicillium roqueforti cultures and from biotransformation of PR toxin". J. Org. Chem. 42 (15): 2632–2634. doi:10.1021/jo00435a023. PMID 874620.
- 1 2 3 4 5 6 Moulé, Y; et al. (1978-04-15). "Inhibition of protein synthesis by PR toxin, a mycotoxin from Penicillium roqueforti". FEBS Letters. 88 (2): 313–323. doi:10.1016/0014-5793(78)80207-5. PMID 648640.
- ↑ Aujard, C.; et al. (1979). "Biochemical effects of PR toxin on rat liver mitochondrial respiration and oxidative phosphorylation". Archives of Biochemistry and Biophysics. 230 (2): 400–411. doi:10.1016/0003-9861(84)90420-x. PMID 6324685.
- ↑ Campbell, J.A.; et al. (1983). "Cheese worker's hypersensitivity pneumonitis". American Review of Respiratory Disease. 127 (4): 495–496. doi:10.1164/arrd.1983.127.4.495. PMID 6838056.
- ↑ Chen, F.C.; et al. (1982). "Acute toxicity of PR toxin, a mycotoxin from Penicillium roqueforti". Toxicon. 20 (2): 433–441. doi:10.1016/0041-0101(82)90006-x. PMID 7080052.