




WW domain-containing transcription regulator protein 1 (WWTR1), also known as Transcriptional coactivator with PDZ-binding motif (TAZ), is a protein that in humans is encoded by the WWTR1 gene. WWTR1 acts as a transcriptional coregulator and has no effect on transcription alone. When in complex with transcription factor binding partners, WWTR1 helps promote gene expression in pathways associated with development, cell growth and survival, and inhibiting apoptosis.[6] Aberrant WWTR1 function has been implicated for its role in driving cancers.[7][8][9] WWTR1 is often referred to as TAZ due to its initial characterization with the name TAZ. However, WWTR1 (TAZ) is not to be confused with the protein tafazzin, which originally held the official gene symbol TAZ, and is now TAFAZZIN.
Structure

WWTR1 contains a proline rich region, TEAD binding motif, WW domain, coiled coil region, and a transactivation domain (TAD) containing the PDZ domain-binding motif. WWTR1 (TAZ) lacks a DNA binding domain so it can not directly drive transcription. WWTR1 exhibits conserved structural homology with another transcriptional coregulator, yes-associated protein 1 (YAP).[6] Both YAP and TAZ are able to form homodimers and heterodimers with each other through interactions at the coil coil domain.[11] YAP and TAZ cooperate with transcription factors to promote tissue formation. WWTR1 (TAZ) interacts with a variety of transcriptional partners, including the four TEA domain family members (TEAD1/2/3/4) through the TEAD-binding motif and several other factors containing the PPXY motif, which consists of a Proline-Proline-X (any amino acid)-Tyrosine sequence. Examples of such partners include Runx/PEBP2, AP2, C/EBP, c-Jun, Krox-20, Krox-24, MEF2B, NF-E2, Oct-4 and p73, which interact with WWTR1 via the WW domain.[6] The transactivation domain at the C-terminal end (amino acids 165–395) was shown to be important in producing transcriptional effects.[6]
Function
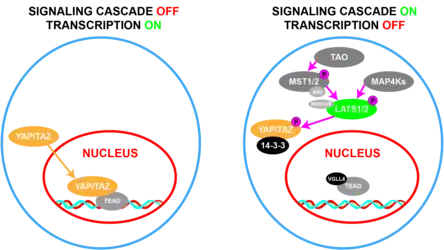
WWTR1 (TAZ) plays an important role in embryogenesis and development,[12][13] which include regulation of organ size,[14][15][16] stem cell renewal,[17] tissue regeneration,[18][17] osteogenesis,[19] and angiogenesis.[20] These functions are effected through coactivation of transcription factors that promote cell growth, migration, and differentiation,[14][15][16] such as the four members of the TEAD transcription factor family, Paired box gene 3 (PAX3), and Runt related transcription factors (RUNX1/)2).[10] The proliferative functions of WWTR1 (TAZ) and its paralog, YAP, are restricted by the Hippo signaling pathway.[21][22][23] This suppressive pathway consists of a kinase signaling cascade, the core of which is made up of the serine-threonine kinases, STK3/MST2 and STK4/MST1, which when active and complexed with the regulatory protein, SAV1, will phosphorylate and activate the LATS1/2 kinases, which in complex with the regulatory protein, MOB1, phosphorylate and downstream inactivate YAP/TAZ.[13][14][24] In this way, Hippo activation arrests cell growth by decreasing proliferative gene expression, leading to decreased cell death by ferroptosis[25][26] and increased cell death by apoptosis.[13][14]
Functional Redundancy with Yes Associated Protein (YAP)
Similarities
WWTR1 (TAZ) has a similar structural sequence and binding motifs to yes-associated protein 1 (YAP).[10] YAP and TAZ are often considered functionally redundant in existing literature.[10] Both play roles in organ size development as well as cell migration, wound healing, angiogenesis, and metabolism, particularly in lipogenesis.[10][27] Inactivation of YAP and TAZ occurs through phosphorylation by kinases in the Hippo pathway, namely LATS1 and LATS2.[10] This recruits the binding of the regulatory protein, 14-3-3, which prevents YAP/TAZ from localizing to the nucleus and marks it for ubiquitination, which allows it to be recognized for subsequent degradation by proteasomes.[10]
Differences
TAZ is able to form both heterodimers and heterotetramers with TEADs to initiate transcription (TAZ-TEAD and TAZ-TEAD-TAZ-TEAD), while YAP is only able to form YAP-TEAD heterodimers.[10] These differences impart unique functions to TAZ, such as in the regulation of adipocyte differentiation through interactions with the peroxisome proliferator-activated receptor (PPARγ), as well as osteogenesis through transcriptional coactivation of bone-specific transcription factors, such as RUNX2 (also known as Cbfa1.)[10] Additionally, TAZ independently interacts with Nuclear factor of activated T-cells 5 (NFATC5) in order to repress transcription in renal cells that are undergoing osmotic stress.[10] Both YAP and TAZ associate with Mothers against decapentaplegic family transcription factors (SMAD) complexes to promote TGF-beta signaling and drive differentiation and development, but upregulation of only TAZ occurs upon transduction of this cascade.[10] TAZ is only able to complex with SMAD2, SMAD3, or SMAD4 to promote nuclear shuttling and transcription, but YAP can also interact with SMAD1 and SMAD7 in addition.[10] In vivo murine studies have demonstrated that animals lacking functional TAZ are more viable than animals lacking YAP expression.[10] In contrast, silencing of YAP contributed to a more dramatic effect on cell expansion, glucose uptake, and cell cycle arrest than TAZ.[10] When assayed in non-small-cell lung cancer (NSCLC) cell lines, WWTR1 maintained the extracellular matrix (ECM) organization and adhesion, and controlled migration more than YAP, which more closely regulated cell division and cell cycle progression genes.[10]
WWTR1 Protein Interactions
_-_Protein_Data_Bank.png.webp)
| Protein Interaction Partner | Functional Effects |
|---|---|
| AMOT, Angiomotin | Binding sequesters YAP/TAZ in the cytoplasm, inhibiting their function[11] |
| AP-1, Activator protein 1 | Promoting trancsription[10] |
| ASPP2, Apoptosis-stimulating protein of p53 | Promotes dephosphorylation and stabilization of WWTR1 (TAZ)[11] |
| β-catenin | Recruits a destruction complex that inactivates YAP/TAZ[11] |
| LATS1/LATS2, Large tumor suppressor kinases | Phosphorylation of WWTR1, marking it for ubiquination[10] |
| NFATC5, Nuclear factor of activated T-cells 5 | Represses transcription in renal cells undergoing osmotic stress[10] |
| PF, Parafibromin | Stimulates WWTR1 (TAZ) function[11] |
| PAX3, Paired box gene 3 | Promoting transcription[10] |
| PAX8 (Paired box gene 8) and NKX2-4 (NK homeobox)[24] | Coactivation of transcription factors involved in thyroid regulation[24] |
| PRRG4, Proline Rich And Gla Domain 4[24] | Suppressing transcription[10] |
| RUNX1/2, Runt related transcription factors | Promotes transcription[10] |
| SMAD2/3/4, Mothers against decapentaplegic family transcription factors | Nuclear shuttling; promoting transcription[10] |
| STAT1, Signal transducer and activator of transcription | Inhibiting STAT1/2 dimerization in metabolism[10] |
| TEAD1, TEA domain family member[24] | Transcriptional activation[10] |
| TEAD2, TEA domain family member,[24] | |
| TEAD3, TEA domain family member,[24] | |
| TEAD4, TEA domain family member[24] | |
| YAP1, Yes-associated protein 1 | Dimerization dependent transcriptional regulation[10] |
| YWHAE (14-3-3), Tyrosine 3-Monooxygenase/Tryptophan 5-Monooxygenase Activation Protein Epsilon[24] | Restricts WWTR1 translocation to the nucleus[10] |
| ZO-2, Tight junction protein 2 | Localizes YAP/TAZ to the nucleus for increased activity[11] |
Clinical Significance
Roles in Diseases
WWTR1 has been implicated in many inflammatory diseases, including cancers.
| Disease | Clinical Significance of WWTR1 |
|---|---|
| Cancer | Associated with metastasis and poor survival prognosis across many cancer types[9] |
| Steatohepatitis | Overexpression of WWTR1 progresses simple steatosis to steatohepatitis by promoting fibrosis[28] |
| Atherosclerosis | Drives excessive endothelial cell proliferation and inflammation[29] |
| Sjogren Syndrome | Decreased WWTR1 localization to the nucleus results in lack of functional salivary/lacrimal gland development[13] |
| Hypertension | Activation of YAP/TAZ promotes glutamine metabolism and increases pulmonary blood pressure[30] |
| Psoriasis | YAP/TAZ activation drive pathologic angiogenesis and inflammation associated with chronic skin disorders[31] |
| Atopic Dermatitis | |
| Rosacea | |
| Chronic Urticaria |
Cancers
WWTR1 (TAZ) is implicated a wide variety of cancers including melanoma, head and neck squamous cell carcinoma, breast cancer, non-small cell lung cancer, and others due to its high gene and histological expression, as well as correlation with increased metastasis and poorer survival in animal studies and patient data.[9] Along with the structurally similar co-regulator YAP, many studies have described their role in promoting oncogenesis, altering neoplastic metabolism, and generating resistance to therapeutic intervention.[8][9][32][33] In particular, TAZ overexpression conferred resistance to cisplatin chemotherapy as well as immunotherapy treatment with a PD-1 antibody.[32]

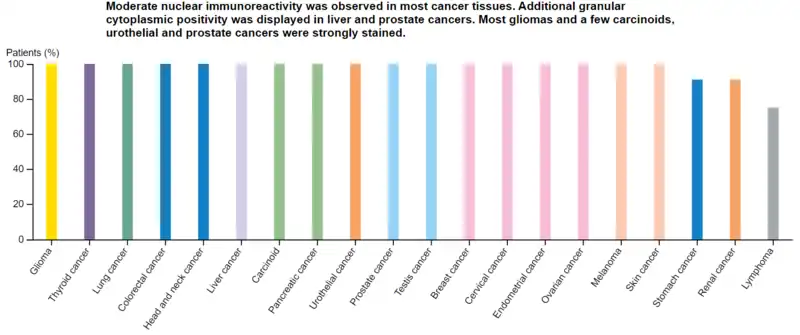
| WWTR1 Protein Expression By Cancer Type (from Protein Atlas) | |||
|---|---|---|---|
| Cancer Type | # Samples with Medium/High Expression | Total # Patient Samples | % Patient Samples with Medium/High Expression |
| Glioma | 11 | 11 | 100% |
| Thyroid | 4 | 4 | 100% |
| Lung | 12 | 12 | 100% |
| Colorectal | 11 | 11 | 100% |
| Head and Neck | 4 | 4 | 100% |
| Liver | 12 | 12 | 100% |
| Carcinoid | 4 | 4 | 100% |
| Pancreatic | 9 | 9 | 100% |
| Urothelial | 9 | 9 | 100% |
| Prostate | 10 | 10 | 100% |
| Testis | 11 | 11 | 100% |
| Breast | 12 | 12 | 100% |
| Cervical | 9 | 9 | 100% |
| Endometrial | 10 | 10 | 100% |
| Ovarian | 11 | 11 | 100% |
| Melanoma | 12 | 12 | 100% |
| Skin | 11 | 11 | 100% |
| Stomach | 10 | 11 | 90.1% |
| Renal | 10 | 11 | 90.1% |
| Lymphoma | 9 | 12 | 75% |
Associated Therapeutics
YAP and TAZ function have been targeted in several therapeutic methods in the treatment of cancers.
The Hippo signaling agonist, C19, increases the phosphorylation of MST1/2 and LATS1/2, resulting in more downstream inactivation of YAP/TAZ. Modulating extracellular matrix stiffness and tension using thiazovivin, cucurbitacin I, dasatinib, fluvastatin, and pazopanib, exhibited positive results in breast cancer cell lines by preventing YAP/WWTR1 translocation to the nucleus.[34] Endogenous hormonal factors that are synthesized for normal physiological functions such as epinephrine and glucagon have also been demonstrated to have similar inhibitory effects on YAP/TAZ function by promoting Hippo pathway activation.[34] The class of cholesterol inhibitors, statins, was shown to inhibit the Rho family of GTP-ases (Rho-GTPase), which are enzymes that signal for upstream inhibition of the Hippo pathway, and exhibited similar effects in attenuating growth of breast cancer and human lung adenocarcinoma cells.[34] Statins inhibit 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase (HMG-CoA reductase), which is the precursor to mevalonate in the mevalonate pathway that synthesizes the lipid building blocks that form cholesterols and the lipid chains responsible for anchoring Rho-GTPases to the cell membrane.[9] The Rho-GTPase, Ras Family Homolog A (RhoA), is activated by prenlylation (the posttranslational modification through addition of hydrophobic groups), and is responsible in part for modulating cytoskeletal elements that reduce Hippo pathway activity.[9] By targeting Rho kinases with thiazovivin, or lipid synthesis through the mevalonate pathway, with statins, RhoA is inhibited and increased Hippo kinase activity may limit proliferation driven by YAP/TAZ.[9][34] Tyrosine kinases signal in proliferative pathways, some which promote YAP/TAZ function, such as Src family kinases and includes the Yes tyrosine kinase, which is associated with YAP function. Targeting tyrosine kinases with inhibitors such as dasatinib and pazopanib has shown some effect in cancers.[9]
Inhibition of YAP/TAZ function by targeting their interactions with their transcriptional partners in the TEAD family has also been studied.[34][35] This includes the use of verteporfin, which was investigated in the treatment of skin cancers, particularly melanoma, although it was not taken beyond preclinical studies.[34]
| Drug/Molecule Name | Drug Class | Mechanism of Action |
|---|---|---|
| C19 | Hippo kinase agonist | Increases phosphorylation by MST1/2 and LATS1/2 to inactivate YAP/TAZ and decrease cell proliferation[34] |
| Dihydrexidine | Dopamine agonist | Increases LATS1/2 activity; Decreases YAP/TAZ function and cell proliferation[34] |
| Epinephrine | Hormonal factor | Increases LATS1/2 activity; Decreases YAP/TAZ function and cell proliferation[34] |
| Glucagon | Hormonal factor | |
| Thiazovivin | Rho kinase inhibitor | Inhibits Rho-GTPase; Increases LATS1/2 activity; Decreases YAP/TAZ function and cell proliferation[9][34] |
| Cucurbitacin I | JAK/STAT3 inhibitor | |
| Dasatinib | Tyrosine kinase inhibitor | |
| Fluvastatin | Statin | |
| Pazopanib | Tyrosine kinase inhibitor | |
| Verteporfin | TEAD inhibitor | Inhibits the binding of YAP/TAZ to TEAD family transcription factors; Decreases proliferative transcription and cell proliferation[34] |
References
- ↑ "AlphaFold Protein Structure Database". alphafold.ebi.ac.uk. Retrieved 2022-11-23.
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000018408 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000027803 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- 1 2 3 4 Hong W, Guan KL (September 2012). "The YAP and TAZ transcription co-activators: key downstream effectors of the mammalian Hippo pathway". Seminars in Cell & Developmental Biology. 23 (7): 785–793. doi:10.1016/j.semcdb.2012.05.004. PMC 3459069. PMID 22659496.
- 1 2 3 Moroishi T, Hansen CG, Guan KL (February 2015). "The emerging roles of YAP and TAZ in cancer". Nature Reviews. Cancer. 15 (2): 73–79. doi:10.1038/nrc3876. PMC 4562315. PMID 25592648.
- 1 2 Zhang X, Zhao H, Li Y, Xia D, Yang L, Ma Y, Li H (September 2018). "The role of YAP/TAZ activity in cancer metabolic reprogramming". Molecular Cancer. 17 (1): 134. doi:10.1186/s12943-018-0882-1. PMC 6122186. PMID 30176928.
- 1 2 3 4 5 6 7 8 9 Zanconato F, Cordenonsi M, Piccolo S (June 2016). "YAP/TAZ at the Roots of Cancer". Cancer Cell. 29 (6): 783–803. doi:10.1016/j.ccell.2016.05.005. PMC 6186419. PMID 27300434.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 Reggiani F, Gobbi G, Ciarrocchi A, Sancisi V (February 2021). "YAP and TAZ Are Not Identical Twins". Trends in Biochemical Sciences. 46 (2): 154–168. doi:10.1016/j.tibs.2020.08.012. PMID 32981815. S2CID 222166778.
- 1 2 3 4 5 6 Callus BA, Finch-Edmondson ML, Fletcher S, Wilton SD (February 2019). "YAPping about and not forgetting TAZ". FEBS Letters. 593 (3): 253–276. doi:10.1002/1873-3468.13318. PMID 30570758. S2CID 58578804.
- ↑ Wu Z, Guan KL (January 2021). "Hippo Signaling in Embryogenesis and Development". Trends in Biochemical Sciences. 46 (1): 51–63. doi:10.1016/j.tibs.2020.08.008. PMC 7749079. PMID 32928629.
- 1 2 3 4 Zheng Y, Pan D (August 2019). "The Hippo Signaling Pathway in Development and Disease". Developmental Cell. 50 (3): 264–282. doi:10.1016/j.devcel.2019.06.003. PMC 6748048. PMID 31386861.
- 1 2 3 4 Piccolo S, Dupont S, Cordenonsi M (October 2014). "The biology of YAP/TAZ: hippo signaling and beyond". Physiological Reviews. 94 (4): 1287–1312. doi:10.1152/physrev.00005.2014. PMID 25287865.
- 1 2 Pocaterra A, Romani P, Dupont S (January 2020). "YAP/TAZ functions and their regulation at a glance". Journal of Cell Science. 133 (2): jcs230425. doi:10.1242/jcs.230425. hdl:11577/3317485. PMID 31996398. S2CID 210945848.
- 1 2 Totaro A, Panciera T, Piccolo S (August 2018). "YAP/TAZ upstream signals and downstream responses". Nature Cell Biology. 20 (8): 888–899. doi:10.1038/s41556-018-0142-z. PMC 6186418. PMID 30050119.
- 1 2 Moya IM, Halder G (April 2019). "Hippo-YAP/TAZ signalling in organ regeneration and regenerative medicine". Nature Reviews. Molecular Cell Biology. 20 (4): 211–226. doi:10.1038/s41580-018-0086-y. PMID 30546055. S2CID 54820180.
- ↑ Driskill JH, Pan D (January 2021). "The Hippo Pathway in Liver Homeostasis and Pathophysiology". Annual Review of Pathology. 16: 299–322. doi:10.1146/annurev-pathol-030420-105050. PMC 8594752. PMID 33234023.
- ↑ Kovar H, Bierbaumer L, Radic-Sarikas B (April 2020). "The YAP/TAZ Pathway in Osteogenesis and Bone Sarcoma Pathogenesis". Cells. 9 (4): 972. doi:10.3390/cells9040972. PMC 7227004. PMID 32326412.
- ↑ Boopathy GT, Hong W (2019). "Role of Hippo Pathway-YAP/TAZ Signaling in Angiogenesis". Frontiers in Cell and Developmental Biology. 7: 49. doi:10.3389/fcell.2019.00049. PMC 6468149. PMID 31024911.
- ↑ Heng BC, Zhang X, Aubel D, Bai Y, Li X, Wei Y, et al. (January 2021). "An overview of signaling pathways regulating YAP/TAZ activity". Cellular and Molecular Life Sciences. 78 (2): 497–512. doi:10.1007/s00018-020-03579-8. PMID 32748155. S2CID 220930261.
- ↑ Ma S, Meng Z, Chen R, Guan KL (June 2019). "The Hippo Pathway: Biology and Pathophysiology". Annual Review of Biochemistry. 88: 577–604. doi:10.1146/annurev-biochem-013118-111829. PMID 30566373. S2CID 58642425.
- ↑ Meng Z, Moroishi T, Guan KL (January 2016). "Mechanisms of Hippo pathway regulation". Genes & Development. 30 (1): 1–17. doi:10.1101/gad.274027.115. PMC 4701972. PMID 26728553.
- 1 2 3 4 5 6 7 8 9 10 11 "UniProt - Q9GZV5 · WWTR1_HUMAN". www.uniprot.org. Retrieved 2022-11-17.
- ↑ Sun T, Chi JT (May 2021). "Regulation of ferroptosis in cancer cells by YAP/TAZ and Hippo pathways: The therapeutic implications". Genes & Diseases. 8 (3): 241–249. doi:10.1016/j.gendis.2020.05.004. PMC 8093643. PMID 33997171.
- ↑ Dai C, Chen X, Li J, Comish P, Kang R, Tang D (September 2020). "Transcription factors in ferroptotic cell death". Cancer Gene Therapy. 27 (9): 645–656. doi:10.1038/s41417-020-0170-2. PMID 32123318. S2CID 211728890.
- ↑ Koo JH, Guan KL (August 2018). "Interplay between YAP/TAZ and Metabolism". Cell Metabolism. 28 (2): 196–206. doi:10.1016/j.cmet.2018.07.010. PMID 30089241. S2CID 51939438.
- ↑ Kuchay MS, Choudhary NS, Mishra SK (2020-11-01). "Pathophysiological mechanisms underlying MAFLD". Diabetes & Metabolic Syndrome. 14 (6): 1875–1887. doi:10.1016/j.dsx.2020.09.026. PMID 32998095. S2CID 222166643.
- ↑ Niu N, Xu S, Xu Y, Little PJ, Jin ZG (April 2019). "Targeting Mechanosensitive Transcription Factors in Atherosclerosis". Trends in Pharmacological Sciences. 40 (4): 253–266. doi:10.1016/j.tips.2019.02.004. PMC 6433497. PMID 30826122.
- ↑ Zhang X, Zhao H, Li Y, Xia D, Yang L, Ma Y, Li H (September 2018). "The role of YAP/TAZ activity in cancer metabolic reprogramming". Molecular Cancer. 17 (1): 134. doi:10.1186/s12943-018-0882-1. PMC 6122186. PMID 30176928.
- ↑ Lee HJ, Hong YJ, Kim M (November 2021). "Angiogenesis in Chronic Inflammatory Skin Disorders". International Journal of Molecular Sciences. 22 (21): 12035. doi:10.3390/ijms222112035. PMC 8584589. PMID 34769465.
- 1 2 Thompson BJ (May 2020). "YAP/TAZ: Drivers of Tumor Growth, Metastasis, and Resistance to Therapy". BioEssays. 42 (5): e1900162. doi:10.1002/bies.201900162. hdl:1885/211659. PMID 32128850. S2CID 212405819.
- ↑ Zanconato F, Cordenonsi M, Piccolo S (August 2019). "YAP and TAZ: a signalling hub of the tumour microenvironment". Nature Reviews. Cancer. 19 (8): 454–464. doi:10.1038/s41568-019-0168-y. PMID 31270418. S2CID 195791034.
- 1 2 3 4 5 6 7 8 9 10 11 Andl T, Zhou L, Yang K, Kadekaro AL, Zhang Y (June 2017). "YAP and WWTR1: New targets for skin cancer treatment". Cancer Letters. 396: 30–41. doi:10.1016/j.canlet.2017.03.001. PMID 28279717.
- ↑ Pobbati AV, Hong W (2020). "A combat with the YAP/TAZ-TEAD oncoproteins for cancer therapy". Theranostics. 10 (8): 3622–3635. doi:10.7150/thno.40889. PMC 7069086. PMID 32206112.