光合作用中心
光合作用中心(英語:)是數种色素、蛋白質亞基和其他辅助因子的复合體,是執行光反應的場所。光反應利用色素分子捕捉光子的能量,并将其变成可用的形式,於碳反應使用。
所有绿色植物、藻类和许多细菌的光合細胞中都有光合作用中心。目前公認的中心有兩種——I型(主要色素為P700)和II型(主要色素為P680)。在绿色植物和藻类細胞中,兩種中心與更多補光複合物(light-harvesting complex,可收集光能,卻不產生光化學反應的色素分子複合體)組成更大、更複雜的功能單元,稱為光系統。红假单胞菌的反应中心目前被研究的最徹底,是第一个已知结构的光合作用中心,其蛋白質結構較植物的單純。[1]
電子轉移與光能轉換
一旦光能被色素分子直接吸收,或通过共振轉移由周围的補光複合物传给色素,發色團(發生光化學反應的色素中因照光產生變化的部位)的電子會被激發,攜帶高能量並脫離原子核靜電力吸引进入电子传递链。電子傳遞鍊中的分子(即電子受體,绿色植物中常見的種類有脫鎂葉綠素、醌、质体醌、细胞色素bf和铁氧还蛋白)一旦接收這些高能電子,就會再釋放一個電子給構成傳遞鍊的下一個原子,透過這種方式將能量最終传递给質子供体如H2O,从中提取电子和质子並釋放氧氣。
質子供體釋放的質子將NADP+還原成NADPH 。电子通过电子传递链也导致质子(氢离子)从叶绿体的基质泵入類囊體内,从而在类囊体膜产生质子梯度,可用于合成ATP(能量貨幣)。ATP和NADPH都可以被用于卡尔文循环(最主要的碳反應形式)。
分类
I型中心可在绿硫细菌、太陽桿菌和植物-蓝藻的光系統I中发现,使用铁硫簇作为电子受体;II型中心則可见于绿弯菌、紫细菌和植物-蓝藻的光系統II,使用醌类作為電子受體。具有相同中心的生物具有共同的祖先。[2][3]
细菌的中心
细菌的光合作用中心一直是了解生物如何將光能轉化為化學能的重要模型,本节讨论在紫細菌中发现的II型系统。1960年代,Roderick Clayton是第一个从紫細菌中纯化出中心复合物的人。第一个晶体结构(右下图)在1984年由哈特穆特·米歇爾 、約翰·戴森霍費爾和羅伯特·胡貝爾[4]确定,這也是第一個被確定的膜蛋白复合物三维晶体结构。細菌的II型系统與植物-藍藻葉綠體膜上的光系統II類似,兩者可能有共同的祖先。
结构
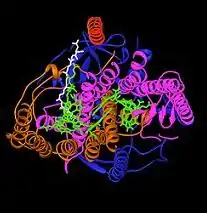
光合細菌的細胞膜向內凹陷,包覆可行光合作用的色素(通常是菌綠素),形成類似藍藻類囊體的囊狀構造,中心自然也位於其中。
細菌的中心中,可行光合作用的色素包含菌绿素b(BChl-b)和脫鎂菌绿素(BPh)b。BChl类似藍藻和植物的叶绿素,但由于微小的结构差异,其主要吸收的光屬於紅外線(葉綠素可吸收可見光),其波长长达1000nm。 Bph与BChl具有相同的结构,但中心镁离子被两个质子取代,导致其吸光度最大偏移且氧化还原电位降低。不同種細菌的BChl-b和BPh可能略有不同,导致不同種細菌可吸收的光譜波段可能不一樣。
細菌的光合作用中心包含四種基本的蛋白質亞基:
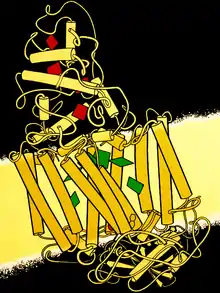
机制
細菌的光反應始於光被位于膜的外延侧附近的两个BChl分子吸收时。这对菌绿素分子,通常被称为 「特殊对」,在870nm或960nm处(取決於物種)吸收光子,因此被称为P870(Rhodobacter sphaeroides菌)或P960(Blastochloris viridis菌),其中P代表色素。BChl吸收了一个光子,就会喷出一个电子,这个电子通过另一个BChl分子转移到L亞基的BPh。电荷轉移造成BChl帶正電(記為P+或P960+),BPh帶負電(記為BPh-)。此步驟发生在10皮秒內。[6]
在这种状态下,P+和BPh-上的电荷可能產生鍵結,放出熵而導致浪费,中心的構造有助于防止这种情况。首先,与反应中心的另外两个氧化还原反应相比,电子从BPh-转移到P+的过程相对缓慢,這兩個較快的反应涉及电子从BPh-(BPh-被氧化为BPh)转移到电子受体醌(QA),以及电子从中心上方的细胞色素亞基的血红素转移到P960+(P960+被还原为P960)。
停留在紧密结合的醌分子QA上的高能电子被转移到一个可交换的醌分子QB上。这个分子与蛋白质松散地联系在一起,相当容易分离。将QB完全还原为QH2需要两个电子,在此过程中从细胞质中吸收两个质子。被还原的醌QH2穿過膜扩散到另一个蛋白复合物(细胞色素bc1-复合物),在那里被氧化。在这个过程中,QH2的还原力被用来将质子穿过膜到周质空间。然后,来自细胞色素bc1-复合物的电子通过周质中的可溶性细胞色素c中间物,即细胞色素c2,转移到细胞色素亚基。
蓝藻和植物的中心
藍藻是綠色植物細胞中叶绿体的前身,具有两种中心系统——光系统II和光系统I,结合这两个系统可以产生氧气。
有氧光合作用
1772年,化学家普里斯特利进行了一系列与氧氣和二氧化碳有关的实验。如果將一隻老鼠與燃烧的蜡烛同置於密閉的玻璃罐中,老鼠在蜡烛熄灭后不久就會死亡;然而,如果以綠色植物取代老鼠,並以陽光照射植物,植物不僅不會在蠟燭燃盡後死亡,甚至還能繼續生長。在近一步研究之後,普里斯特利證明植物在吸收啤酒發酵、蠟燭燃燒以及動物呼吸時產生的「固定氣體」(即二氧化碳)後可以製造能維持動物呼吸、使燃燒更劇烈的「活命氣體」(即氧氣)。普里斯特利在著作中以燃素說的觀點解釋,將氧氣稱為「脫去燃素的氣體」。普利斯特里的观察是最早证明光合作用反应中心活动的实验。
1779 年,詹·英格豪斯在實驗中将绿色植物浸在透明水箱中,每当植物暴露在光线下时,许多气泡从叶子表面浮起。英格豪斯收集了植物放出的气体,并将一个燃烧的锥子放入气体样本中,让它重新点燃,证明植物會放出氧气,即普里斯特利所謂「脫去燃素的氣體」。
1932年,罗伯特‧爱默生(Robert Emerson)和他的学生William Arnold使用重复闪光技术精确测量了小球藻中叶绿素所释放的少量氧气,证明了光合作用单元的存在。Hans Gaffron和Kurt Wohl后来对该实验进行了解释,意识到光合单元在製造氧氣的同時將吸收的光能轉移了。[7]這一轉移能量的過程發生在光系統II的中心,存在於蓝藻、海藻和绿色植物中。[8]
光系统II
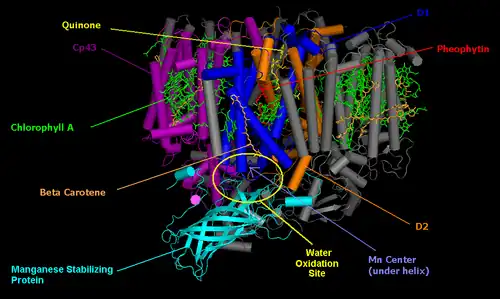
光系统II位於叶绿体膜上,[9]是在光反應中产生两个电子的光系统,最终在鐵氧化蛋白-NADP+还原酶中还原NADP+。光系统II的结构与细菌的光合作用中心非常相似,理论上它们具有共同的祖先。
光系统II的核心由两个亞基D1和D2組成,类似细菌中心的L和M亚基。光系统II比细菌的中心多了许多额外的蛋白質亚基,这些亚基結合额外的叶绿素以提高效率。
光系统II催化的总反应为:
- 2Q + 2H2O + hν → O2 + 2QH2
Q代表质体醌(Q的氧化形式),QH2代表質體酚(Q的还原形式),还原醌的过程与在细菌中心发生的过程相同。光系统II透过光分解作用氧化水以获得电子,產生副產品氧氣,為現今大氣層主要的氧氣來源。绿色植物中的氧气来自水的事實最早是由马丁·大卫·卡门(Martin David Kamen)推断出来的。他使用稳定的氧同位素18O来追踪从水到氧氣分子的反應路径,该反应由光系统II中含四个锰离子的中心催化。
植物的光反應始于一对被称为P680[10]的叶绿素分子激发,与细菌的反應類似。由于植物的特殊對色素為叶绿素a,而不像細菌是菌绿素,光系统II吸收的光波长较細菌的中心短。当P680吸收一個光子后,就會放出一個高能电子給附近的脱镁叶绿素分子,使P680帶正電,此過程被稱為光誘導電荷分離(photoinduced charge separation)。电子从叶绿素分子中穿过两个質體醌分子,第一个是紧密结合的,第二个是松散结合的,这种电子流与细菌反应中心的电子流相似,需要两个电子将松散结合的質體醌分子完全还原成QH2,同时也需要吸收两个质子。
P680在吸收光子→放出电子→帶正电之後,將成為一种非常强的高能氧化剂P680+。它将其能量传递给结合在锰中心(由四个锰离子、一个钙离子、一个氯离子和一个酪氨酸残基組成)正下方的水分子,並氧化它們,使两个水分子形成一个氧分子。到目前为止,這種分水催化中心还没有被任何人造的催化剂所复制。
氧化水後可以得到四个电子,為将两个Q分子还原为QH2的电子源。與细菌不同,在细菌的中心,還原Q分子的电子是从细胞色素亚基中的还原复合血红素基团或水溶性细胞色素-c蛋白中获得。
光系統 I
在电子离开光系统II后,會被转移到细胞色素b6f复合物,然后转移到质体藍素(一种蓝色铜蛋白和电子载体)。质体蓝素复合物携带的电子将中和下一个中心——光系統 I中的电子對。
与光系统II和细菌中心一样,光系統I中一对叶绿素a分子启动了光诱导电荷分离。这对分子被称为P700,位於蛋白質的中心位置,其中700指的是叶绿素分子最大吸收光的波长。一旦光诱导的电荷分离开始,电子就会沿着一条路径,穿过位于P700正上方的叶绿素α分子,穿过位于正上方的醌分子,穿过三个4Fe-4S团簇,最后到达一个可互换的铁氧还蛋白复合物。[11]铁氧还蛋白是一种可溶性蛋白质,含有一个由四个半胱氨酸残基协调的2Fe-2S簇。高能的P700+上的正电荷被来自质体藍素的电子转移所中和,质体藍素获得的能量最终用于将QH2转化为Q。
光系统I催化的整体反应是:
- Pc(Cu+)+Fd[ox]+hν → Pc(Cu2+)+Fd[red]
光系统I和II之间的合作产生了从H2O到NADP+的电子和质子流,产生了葡萄糖合成所需的NADPH。这一途径被称为「Z方案」(Z-scheme),因为从H2O经P680和P700到NADP+的氧化还原图类似于字母Z。[12]
外部链接
- . 伊利諾大學厄巴納-香檳分校. [2022-01-10]. (原始内容存档于2009-12-08).
參考
- . New York: WH Freeman. 2002 [2022-01-10]. (原始内容存档于2010-05-31).
- Sadekar, S; Raymond, J; Blankenship, RE. . Molecular Biology and Evolution. November 2006, 23 (11): 2001–7. PMID 16887904. doi:10.1093/molbev/msl079.
- . Photosynthesis Research. October 2018, 138 (1): 11–37. OSTI 1494566. PMID 29603081. doi:10.1007/s11120-018-0503-2.
- . Journal of Molecular Biology. December 1984, 180 (2): 385–98. PMID 6392571. doi:10.1016/S0022-2836(84)80011-X.
- Photosynthetic reaction centers of purple bacteria 的存檔,存档日期2006-05-14. (2 February 1999). Retrieved Feb 28, 2010.
- 5th. New York: WH Freeman. 2002 [2022-01-10]. (原始内容存档于2010-05-31).
- . London: Taylor & Francis. [2022-01-10]. ISBN 978-0-7484-0821-4. (原始内容存档于2014-05-24).
- Kaiser, Gary E. . 24 February 2003. (原始内容存档于4 May 2006).
- . Ultranet Biology. (原始内容存档于3 August 2003).
- 5th. New York: WH Freeman. 2002 [2022-01-10]. (原始内容存档于2010-05-31).
- Jagannathan, Bharat; Golbeck, John. Schaechter , 编. 3rd. 2009: 325–341. ISBN 978-0-12-373944-5. doi:10.1016/B978-012373944-5.00352-7.
- Govindjee, Rajni. . University of Illinois at Urbana-Champaign. [2022-01-10]. (原始内容存档于2014-06-25).