哈采格島
哈采格島(英文:),或譯哈提格島,為白堊紀晚期(森諾曼期至马斯特里赫特期)一座位於特提斯洋的已消失島嶼[1],位置相當於現今羅馬尼亞胡內多阿拉縣的哈采格[2]。哈采格島的形成肇因於白堊紀晚期非洲板塊與歐亞大陸板塊碰撞,阿爾卑斯造山運動所引發的構造抬升[3]。 在哈采格島上,發現了許多體型特別小的恐龍化石[4],匈牙利古生物學家弗朗茨·諾普札認為島上的資源有限,造成這些恐龍隨著世代逐漸演化出較小的體型。這個島嶼侏儒化理論(或是福斯特法則)如今已被廣泛接受。

地理學
目前普遍認為马斯特里赫特期的哈采格島面積大約落在80,000平方(31,000平方英里),與現今的伊斯帕尼奧拉島大小近似;島嶼的位置則坐落於赤道帶內,北緯27度線上[1]。
哈采格島距離最近的大陸距離至少有200(120英里),西北方則鄰近一座島嶼(對應於現今的波希米亞山)、西南方鄰近另一座島嶼(對應於巴爾幹-洛多皮地塊,包括現今的洛多皮山脈地區)、西方鄰近於對應現今伊比利半島的大型島嶼。距離哈采格島最近的大陸為島嶼西方的奧地利-阿爾卑斯地區與島嶼南方的亞得里亞海地區[1]。
島嶼與大陸周遭大多被淺海圍繞,然而哈采格島卻是位於盆地之中,被深海所環繞[1]。
氣候與環境學
於马斯特里赫特期時,哈采格島屬亞熱帶氣候,平均氣溫約為20—25 °C(68—77 °F)[1],具有乾季與旱季。儘管如此,碳同位素分析島上的植披仍然以熱帶植物為主;原因推測與現今能存活於季風氣候的熱帶植物類似,只要一整年接受到的水分充足,即使面臨乾季這些植物仍然能夠存活。另外哈采格島上也具有大量交織的河流與湖泊,能提供這些植物生存所需的水分。哈采格島較早期的地層中主要為火山沉積物,然後較晚期則無,顯示島上的火山運動很早就已停止[1]。
古動物群

約9種恐龍及數種翼龍被認為是哈采格島上的特有種,這些生存於島上的物種受到島嶼效應的影響,與鄰近大陸的近親在外型與行為上呈現出不同程度的差異。其中包括了島嶼侏儒化,例如泰坦巨龍類的馬扎爾龍,體重僅有900(2,000英磅)[5][6],而生存於大陸的泰坦巨龍巴塔哥巨龍體重則可達69公噸(76英噸)。除此之外,有些翼龍如哈特茲哥翼龍則有島嶼巨型化的趨勢,成為了目前已知體型最大的翼龍之一。如同現存生存於島嶼的鳥類一樣,粗壯巴拉烏爾龍也同樣失去了飛行能力。這樣的特例也導致巴拉烏爾龍最初在分類上的誤判,在最初被發現時,牠們由於具有較為粗短的前肢,被分類至恐爪龍下目下;然而近年來透過種系發生學研究分析結果顯示,巴拉烏爾龍其實是鳥翼類的基群物種。島上的哺乳動物幾乎全部皆為多瘤齒獸目科蓋奧農獸科的物種,為食蟲動物[7],另外也有發現真獸下綱物種的化石[8]。
發現於哈采格島上的動物包括:
- 先驅異鄉鱷 Allodaposuchus precedens
- 合齒劍鱷 Sabresuchus sympiestodon
- 巴爾巴特齒獸屬 Barbatodon sp.
- 昂氏科蓋奧農獸 Kogaionon ungureanui
- 曲顱統治者獸 Litovoi tholocephalos
- 怪物哈采格翼龍 Hatzegopteryx thambema
- 原始凹齒龍 Rhabdodon priscus
- 強壯查摩西斯龍 Zalmoxes robustus
- 外西凡尼亞沼澤龍 Telmatosaurus transylvanicus
- 外西凡尼亞鴕龍 Struthiosaurus transylvanicus
- 達契亞馬扎爾龍 Magyarosaurus dacus
- 納拉次沼澤巨龍 Paludititan nalatzensis
- 諾氏沼澤鳥龍 Elopteryx nopcsai
- 匈牙利斑龍 Megalosaurus hungaricus?
- 粗壯巴拉烏爾龍 Balaur bondoc
- 德氏重腿龍 Bradycneme draculae
- 朗根多夫歐洲神龍翼龍 Eurazhdarcho langendorfensis
- 塔米斯阿爾巴翼龍 Albadraco tharmisensis
- 未命名的神龍翼龍科物種
- 格氏小鱷 Aprosuchus ghirai,目前已知生存年代最接近現代的阿吐波鱷科物種


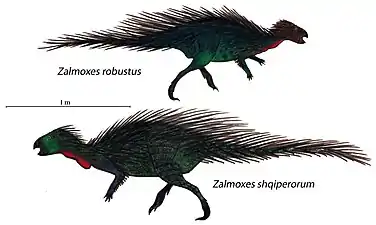 查摩西斯龍屬物種
查摩西斯龍屬物種
參見
- 哈采格恐龍地質公園
- 羅馬尼亞的恐龍
- 弗朗茨·諾普札·馮·法西歐-錫爾瓦什
- 島嶼侏儒化
參考文獻
- Benton, M.J.; Csiki, Z.; Grigorescu, D.; Redelstorff, R.; Sander, P.M.; Stein, K.; Weishampel, D.B. (PDF). Palaeogeography, Palaeoclimatology, Palaeoecology. 2010, 293 (3-4): 438–454. (原始内容 (PDF)存档于2011-07-10).
- Csikia, Z.; M. J. Bentonb. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2010, 293 (3–4): 265–270. Bibcode:2010PPP...293..265C. doi:10.1016/j.palaeo.2010.05.032.
- Tom Baird and Richard Conium. . Paleobiology and Biodiversity Research Group, Department of Earth Sciences, University of Bristol. [March 16, 2020]. (原始内容存档于2021-04-21).
It was formed by uplift caused by the closing of the Tethys Ocean by the collision of the African and Eurasian plates
- Grigorescu, D. . Acta Palaeontologica Romaniae. 2005, 5: 191–204.
- Carballido, J.L.; Pol, D.; Otero, A.; Cerda, I.A.; Salgado, L.; Garrido, A.C.; Ramezani, J.; Cúneo, N.R.; Krause, J.M. . Proceedings of the Royal Society B: Biological Sciences. 2017, 284 (1860): 20171219. PMC 5563814
 . PMID 28794222. doi:10.1098/rspb.2017.1219.
. PMID 28794222. doi:10.1098/rspb.2017.1219. - Stein, K.; Csiki, Z.; Rogers, K. C.; Weishampel, D. B.; Redelstorff, R.; Carballido, J. L.; Sander, P. M. . Proceedings of the National Academy of Sciences. 30 April 2010, 107 (20): 9258–9263. Bibcode:2010PNAS..107.9258S. PMC 2889090 . PMID 20435913. doi:10.1073/pnas.1000781107 .
- Smith, Thierry; Vlad, Codrea. . PLoS ONE. 2015, 10 (7): e0132550–1–16. doi:10.1371/journal.pone.0132550.
- Codrea, Vlad; Smith, Thierry; Dica, Paul; Folie, Annelise; Garcia, Géraldine; Godefroit, Pascal; Van Itterbeeck, Jimmy. . Comptes Rendus Palevol. 2002, 1 (3): 173–180. ISSN 1631-0683. doi:10.1016/S1631-0683(02)00021-0.