巴哈達龍屬
巴哈達龍(學名:Bajadasaurus,發音:bæ-hæ-dæ-s-ɔːr-ə-s,意為「坡地的蜥蜴」,或譯作巴加達龍或坡地龍)是一屬梁龍超科的蜥腳下目恐龍,生存於白堊紀早期晚貝里亞階至凡藍今階,地理位置相當於現在的阿根廷巴塔哥尼亞北部。模式種兼唯一種前傾棘巴哈達龍(Bajadasaurus pronuspinax)是根據2010年發現的完整頭骨以及部分頸部化石,於2019年發表正式的敘述。[1]巴哈達龍屬於叉龍科,這是一群有著特殊短頸部及小體型的蜥腳類,生存於侏儸紀早期或中期至白堊紀早期結束。
| 巴哈達龍屬 | |
|---|---|
 | |
| 巴哈達龍的骨骼材料 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | †蜥脚形亚目 Sauropodomorpha |
| 下目: | †蜥腳下目 Sauropoda |
| 科: | †叉龙科 Dicraeosauridae |
| 属: | †巴哈達龍屬 Bajadasaurus Gallina et al., 2019 |
| 模式種 | |
| †前傾棘巴哈達龍 Bajadasaurus pronuspinax Gallina et al., 2019 | |
巴哈達龍擁有由頸椎神經棘延長而成的極長棘刺,化石更完整的近親阿馬加龍也有類似的長刺構造。過去阿馬加龍棘刺的功能就經常被拿來討論,隨著巴哈達龍的被敘述,有推論說明棘刺功能是作為對掠食者的被動防禦。頭骨纖細,長著44顆鉛筆狀牙齒,侷限於頜部前端。眼窩可見於頭部上方,有助於覓食時視野朝前,可能形成雙眼視覺。巴哈達龍是在紅坡組沉積岩中發現,當地過去的環境是個辮狀河系統,在那裡巴哈達龍與同為梁龍超科的遠親末梁龍以及其他獸腳類共享棲地。[1]
發現與命名
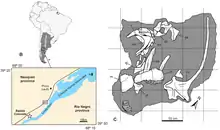
唯一的標本是由阿根廷政府科學機構CONICET的古生物學家所發掘。發現於皮昆留富鎮以南40公里處,靠近利邁河西岸的紅坡(Bajada Colorada)地區。標本最初僅有部分牙齒暴露在外,是由阿根廷古生物學家帕部羅·加里納(Pablo Gallina)所發現。由於當地的化石普遍脆弱,因此標本不是在野外直接挖掘,而是將整塊大岩石與骨頭一同包裹在石膏中搬下來。其後帶到實驗室裡處理岩塊,清修出一隻新屬恐龍的大部分頭骨、前兩節及可能的第五節頸椎。[2][1]發現地屬於紅坡組地層的一部分,是內烏肯盆地中的一系列沉積岩,年代約為下白堊紀的晚貝里亞階至凡藍今階。標本現在在內烏肯省喬昆城的爾尼斯托·巴赫曼古生物博物館展出,編號為MMCh-PV 75。[1]
標本由加里納和同事於2019年正式命名、敘述、發表為新屬、新種-前傾棘巴哈達龍(Bajadasaurus pronuspinax)。屬名取自發現地名Bajada(在西班牙文意為「下坡」),種名由拉丁文的pronus(向前彎曲)和希臘文的spinax(棘刺)組成,意指其頸部長而向前彎曲的神經棘。[1]
敘述

巴哈達龍被分類於蜥腳下目的叉龍科。如同所有蜥腳類,叉龍科是大型四足的草食性動物,具有長頸長尾和相當小的頭部。與其他蜥腳類相比,叉龍科體型偏小,大約等同於現代的亞洲象,其頸部也相對得短。[3][4]雖然長神經棘是叉龍科的普遍特徵,但只有巴哈達龍和近親阿馬加龍有極度加長的棘刺。[1]
頭骨

頭骨包含了大部分頭蓋骨和腦殼、腭骨(palate)的翼骨(pterygoid)、以及下頜和部分的上頜,並且是目前已知最完整的叉龍科頭骨。頭骨中段未保存下來。整體形狀纖巧。[1]
環眶部基本上都有保存下來,除了下眶緣的顴骨。眼眶前緣的淚骨上半部有一道直脊,類似叉龍。脊有小孔洞穿過,不像在叉龍上見到的大孔洞。前額骨組成眼眶的前上角,但前額與眼眶的接面比叉龍或阿馬加龍的都小,骨骼也比後兩者的小更不堅固。眼眶上部由額骨組成,並與後方的頂骨融合成一塊,形成頂骨的大部分後部。俯視額骨側緣呈S形,並從後向前變窄。於是在頂部可見到眼睛的開口,不同於其他叉龍科物種,除了靈武龍。後視額骨形成了上顳孔(頭骨後部的主要孔洞)的一小部分。眼眶後部由眶後骨組成,通常在恐龍中這個骨頭有向後延伸的突起,稱為後突,但在巴哈達龍、叉龍、阿馬加龍中,這個突起退化而變得不明顯。鱗狀骨(形成頭骨的後上角)的向下突起發達,這代表著它與頭部後下部的方軛骨相連,雖然關節本身並未保存下來。這道可能的關節缺乏於梁龍科,且之前也未在叉龍科記錄到。顳顬孔是由鱗狀骨、眶後骨、方軛骨和顴骨圍成的孔洞,是頭骨的另一個主要開口,巴哈達龍的孔狹窄而傾斜。方軛骨形成鈍角包圍著顳顬孔的後下部,與梁龍科的情況不同。[1]

腦殼大部分被其他骨骼覆蓋而遮住,只有枕骨區(後部)露出。枕骨最高處為上枕骨,而巴哈達龍的與下方的後凹外枕骨融合,並形成明顯狹窄的縱脊,稱為頸矢狀嵴。頂骨與枕骨區之間的一對開口-後顳孔朝向頭骨的中心平面延伸至內側,是巴哈達龍的獨有衍徵。枕髁(與第一節頸椎關節連接處)寬而高,其後部表面不比頸部寬,不同於阿馬加龍和叉龍。基蝶骨構成腦殼下部,有一對纖巧的骨質延伸,稱為基蝶突,向前及向下延伸與腭骨的翼骨連接,從而包圍腦殼使之與後者區隔。其中一個自衍徵是這些突起比叉龍或阿馬加龍的還要長且纖細,長度比寬度長了六倍。腭骨唯一保存下來的左右翼骨具平滑的冠狀以與基蝶谷突起連接。[1]
牙齒局限於頜部前端並呈鉛筆狀,窄齒冠幾乎筆直或向內稍微彎曲。上頜部分,只有左上頜骨(上頜最大的骨頭)的前部保存下來。保存了八個齒槽,數量與春雷龍相同,但比叉龍的12個要少。看似完整的一排24顆牙齒與上頜骨分離、在附近找到。此計數與下頜的牙齒計數相對應,下頜牙齒仍固定在左右齒骨(下頜唯一長有牙齒的骨頭)。因此巴哈達龍總共可能有44顆牙齒。齒骨纖細,類似春雷龍但不像叉龍的那麼深。俯視圖中,齒骨未形成梁龍科見到的盒狀口鼻部,但更圓潤呈J形曲線,就像典型的叉龍科。齒骨前端長著鉤狀的下巴,就如同其他鞭尾類所見。下頜後部的隅骨大幅加長並長於上隅骨,不同於梁龍科。[1]
頸椎
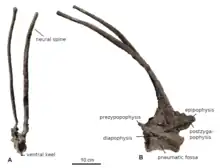
在與頭骨的關節中發現兩個前寰椎(位於第一個頸椎骨和頭骨之間的三角形小骨頭)。寰椎(第一頸椎)只有上部,即寰神經突保存下來,巴哈達龍的是三角形且翅膀狀。樞椎(第二頸椎)幾乎完整,就像叉龍,高度為長度的兩倍,而椎體長度是高度的兩倍。橫關節突(側向突起)小且直接向後,如同春雷龍,而不像叉龍和阿馬加龍的向下。樞椎神經棘狹窄無分叉,垂直向上,不同於其他蜥腳類(自衍徵)。橫截面為三角形,向頂端逐漸變細。[1]
頸部其餘部分只有另一節頸椎保存下來,這節頸椎具有本屬最重要的特徵—極度加長的神經棘,深深地分成左右兩條分叉。長刺狀,長約58公分,使脊椎高度是長度的四倍。在其他所有蜥腳類中,唯有阿馬加龍有相同特徵。但與阿馬加龍不同的是,巴哈達龍的長刺不是向後,而是向前彎曲。底部三角形側扁,沿大部分長邊的橫切面為卵圓形。尖端略變寬,不像阿馬加龍的急遽變細。阿馬加龍的棘刺表面有條紋,代表著活著時包覆著角質鞘。雖然巴哈達龍因化石保存狀態不佳,未找到類似的條紋,但加里納和同事推測,應該也包覆著同樣的角質鞘。然而這個頸椎在頸部的實際位置不得而知,形態上類似叉龍的第五節、短頸潘龍的可能第六節、阿馬加龍的第七節;根據這些比較,它被暫時描述成第五頸椎。椎體長度為高度的兩倍,底側縱脊漸窄;這個脊比其他叉龍科的更寬、更凹。[1]
分類

叉龍科是梁龍超科三個主要的科之一,而梁龍超科是蜥腳下目的主要演化支之一。在梁龍超科中,叉龍科和梁龍科互為姐妹群,而雷巴齊斯龍科則關係較遠。前兩者一同組成鞭尾類,一群以鞭狀尾部得名的類群。[5]加里納和同事2019年的敘述中,將巴哈達龍分類為叉龍科的第七屬。已知叉龍科最早的物種靈武龍生存於侏儸紀早期或中期的中國;[6]三個物種來自侏儸紀晚期:坦尚尼亞的叉龍、阿根廷的短頸潘龍和美國的春雷龍;白堊紀早期的物種包含巴哈達龍、阿馬格巨龍、皮爾馬圖埃龍、阿馬加龍,全部來自阿根廷;另一個來自巴西的未命名物種,暗示著叉龍科可能倖存至白堊紀早期的結束。[7]

加里納和同事的系統發生學分析將巴哈達龍列於叉龍科的中階物種,比春雷龍、靈武龍衍化,但比皮爾馬圖埃龍、阿馬加龍、叉龍、短頸潘龍原始;阿馬格巨龍因位置的不確定從討論中被排除。他們根據六項共有衍徵來支持巴哈達龍屬於叉龍科。而巴哈達龍擁有四個自衍徵,包含:後顳孔向內延伸;基翼骨細長;第二頸椎神經棘垂直;頸椎神經棘加長、向前彎曲。[1]
以下系譜樹取自加里納等人(2019)的研究,顯示叉龍科可能的物種親緣關係:[1]
| 梁龍超科 Diplodocoidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||
古生物學
棘刺功能
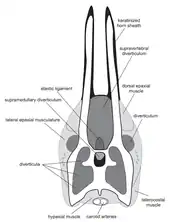
加長和深度分叉的神經棘在叉龍科很常見。在叉龍和短頸潘龍中,它們的神經棘朝前傾斜,但比巴哈達龍的短很多。只有阿馬加龍脊椎有類似的加長情況。阿馬加龍的脊椎引起了關於它們的生前外觀及用途的許多猜測。根據不同學者的假設,它們本來可能支撐著帆狀物或角質鞘,並可能作為展示、防禦、或體溫調節用途。[1]2007年丹妮拉·史瓦茨(Daniela Schwarz)和同事的研究發現,沿叉龍科脊椎的神經棘雙叉形成的空間中,封閉著一個氣囊,即椎上憩室,作為呼吸系統的一部分與肺相連。在叉龍中,這個氣囊佔據神經棘左右兩側之間的整個空間,而在阿馬加龍中,它侷限於神經棘下方三分之一的部分。如表面縱向條紋所示,神經棘上方三分之二的部分可能有角質鞘覆蓋著。[8]
加里納等人(2019)認為這項最具合理性的理論亦適用於巴哈達龍。這些研究人員進一步指出,角質鞘的材質與骨頭相比更能抵抗衝擊所造成的骨折,因此角質鞘可以保護脆弱的脊椎免受傷害。脊椎損害可能是一項嚴重的威脅,因其底下包覆著脊髓。而角質鞘超出骨核帶則進一步加強保護性。史瓦茨和同事在重建阿馬加龍的角質鞘時沒有將它們延伸至其骨核帶,如同許多現代爬蟲類的情況。在一些現代偶蹄目中,角質鞘的長度可達角核帶的兩倍,並發現在精美保存下來的甲龍類北方盾龍其角質鞘延伸了脊椎長度的25%,證實了角質鞘的延伸情況也出現於恐龍中。加里納和同事推測阿馬加龍和巴哈達龍的脊椎可能比骨核帶延長50%左右的長度[1]。它們的角質鞘彎曲能進一步增強抵抗力,就像現代的大角羊一樣。[1][9]
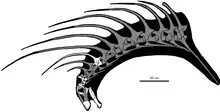
加里納和同事進一步推測,阿馬加龍和巴哈達龍的棘刺都可能用來防禦。由於棘刺向前彎曲,假定第五頸椎的分叉神經棘會達到頭部上方,就像用圍欄一樣來阻止掠食者靠近。同時還假設後方頸椎的棘刺會變得更大。適度的磨損會導致角質鞘尖端折斷,使骨質部分碰觸到外面。阿馬加龍生活在晚於巴哈達龍的1500萬年後,代表著這種加長神經棘是一種持久適用的防禦策略。[1]
古生態學
巴哈達龍出土於紅坡組,是巴塔哥尼亞北部門多薩群(Mendoza Group)露出的地質構造。由紅色和青棕色的砂岩、由細至粗的礫岩、帶紅色的黏土岩以及淺棕色的粉砂岩所組成。這些沉積物由辮狀河堆積,呈現出保存良好的河道與粒徑。該地質構造亦呈現了古土壌。紅坡組覆蓋著昆圖科組(Quintuco Formation)及皮昆留富組(Picún Leufú Formation)。[11][1][12]上部則與阿格里奧組約1.34億年前的不整合面作分隔。[13]巴哈達龍來自構造的模式區-紅坡區。當地還發現另一種蜥腳類-梁龍科的末梁龍,以及其他幾種獸腳類,包含可能的堅尾龍類、阿貝力龍超科和馳龍科。[11][1]
參考資料
- Gallina, P.A.; Apesteguía, S.; Canale, J.I.; Haluza, A. . Scientific Reports. 2019, 9 (1): 1392. Bibcode:2019NatSR...9.1392G. PMC 6362061
 . PMID 30718633. doi:10.1038/s41598-018-37943-3.
. PMID 30718633. doi:10.1038/s41598-018-37943-3. - Yong, Ed. . The Atlantic. 2019-02-05 [2019-02-09]. (原始内容存档于2021-01-21).
- Hallett, M.; Wedel, M. . Baltimore: Johns Hopkins University Press. 2016: 45. ISBN 978-1421420288.
- Paul, G. S. (PDF). The Dinosaur Report. 1994, 8 [2020-08-13]. (原始内容存档 (PDF)于2021-03-16).
- Harris, JD; Dodson, P. . Acta Palaeontologica Polonica. 2004, 49 (2): 197–210.
- Xing Xu; Paul Upchurch; Philip D. Mannion; Paul M. Barrett; Omar R. Regalado-Fernandez; Jinyou Mo; Jinfu Ma; Hongan Liu. . Nature Communications. 2018, 9 (1): Article number 2700. Bibcode:2018NatCo...9.2700X. PMC 6057878 . PMID 30042444. doi:10.1038/s41467-018-05128-1.
- Novas, F.E. . Bloomington: Indiana University Press. 2009: 172–174. ISBN 978-0-253-35289-7.
- Schwarz, D.; Frey, E.; Meyer, C. A. (PDF). Acta Palaeontologica Polonica. 2007, 52 (1) [2020-08-14]. (原始内容存档 (PDF)于2016-11-03).
- Drake, Aaron; Haut Donahue, Tammy L.; Stansloski, Mitchel; Fox, Karen; Wheatley, Benjamin B.; Donahue, Seth W. . Acta Biomaterialia. 2016-10-15, 44: 41–50. ISSN 1742-7061. PMID 27544811. doi:10.1016/j.actbio.2016.08.019.
- Paulina Carabajal, A.; Carballido, J.L.; Currie, P.J. . Journal of Vertebrate Paleontology. 2014, 34 (4): 870–882. doi:10.1080/02724634.2014.838174.
- Gallina, P.A.; Apesteguía, S.Z.; Haluza, A.; Canale, J.I. . PLoS ONE. 2014, 9 (5): e97128. Bibcode:2014PLoSO...997128G. PMC 4020797 . PMID 24828328. doi:10.1371/journal.pone.0097128.
- Leanza, Héctor A.; Hugo, Carlos A. . Publicación Electrónica de la Asociación Paleontológica Argentina. 2001, 7 (1): 116–122 [2020-08-14]. (原始内容存档于2019-02-12).
- Leanza, Héctor A. . Anales de la Academia Nacional de Ciencias Exactas, Físicas y Naturales. 2005, 57: 147–155.