盤足龍屬
盤足龍(屬名:Euhelopus,意為「真正的沼澤腳」)是一屬蜥腳下目多孔椎龍類恐龍,生存於白堊紀早期(1.29至1.13億年前)的中國山東省[1]。它是大型的四足草食性恐龍。不同於一般的蜥腳類,盤足龍的前肢長於後肢。盤足龍的發現在科學上有著重大意義,它是中國第一個進行科學研究的恐龍:1913年首次發現、1922年再次發現、1923年挖掘出土、1929年發表敘述[2]。與許多蜥腳類標本不同的是,盤足龍保存了罕見、相對完整的頭骨[3]。
| 盘足龙属 化石时期:下白堊紀, | |
|---|---|
 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | †蜥脚形亚目 Sauropodomorpha |
| 下目: | †蜥腳下目 Sauropoda |
| 科: | †盘足龙科 Euhelopodidae |
| 属: | †盘足龙属 Euhelopus Wiman, 1929 |
| 模式種 | |
| †師氏盤足龍 Euhelopus zdanskyi Wiman, 1929 | |
發現歷史
最初的發現是由天主教神父R. Mertens發現於1913年,他給德國採礦工程師Gustav Behaghel看了他所挖的一些遺骸,後者並在1916年將三個脊椎交給中國地質調查局局長丁文江(V.K. Ting)。這可能是第一次對中國的恐龍骨骼進行科學研究。藉由另一位天主教神父Alfred Kaschel的幫助之下,約翰·岡納·安德森和譚錫超於1922年11月再次發現挖掘地。1923年3月,奧地利學生奧托·師丹斯基在相距約三公里的地點挖掘了兩個骨骼[2]。

起初於1929年由瑞典古生物學家卡爾·衛曼命名為Helopus(意指沼澤的腳),由希臘文的ἕλος/helos(沼澤)和πούς/pous(腳)組成[4]。名稱指的是發現地的沼澤以及瑞典沼澤靴truga,根據衛曼所說這種靴子類似動物的寬腳[4]。但因這個學名已被一種紅嘴巨鷗Helopus caspius(Wagler, 1832)所使用,所以這個蜥腳類後來在1956年由阿爾弗雷德·羅默(Alfred Sherwood Romer)更名為Euhelopus(真正的沼澤腳)[5]。植物界有一種草的屬名也使用了Euhelopus,但不同界允許相同的屬名,因此Euhelopus依然是有效的。模式種是師氏盤足龍(Helopus zdanskyi/Euhelopus zdanskyi),種名紀念化石挖掘者師丹斯基[4]。

標本編號PMU 24705(之前的PMU R233)根據Wilson & Upchurch描述成為師氏盤足龍的正模標本[1][3]。其包含了師丹斯基所發掘的一副骨骼,由衛曼命名為「Exemplar a」但並未正式歸入模式標本[4]。這也是Mertens所發現的原始骨骼[4]。PMU 24705標本包含了顱骨和下頜,以及部分挖掘的部分骨骼:左鼻骨的延髓部分、部分右顴骨、眶後骨的錐形顴骨突;右方軛骨的背突、左翼骨碎片(另一碎片可能是右夾板骨,但太破碎無法從基部移除)[3]、一系列25節薦前椎和左大腿骨[4]。

第二具骨骼是另一個個體,體型接近於正模標本,由衛曼標註為「Exemplar b」[4]。該標本由Wilson & Upchurch歸於盤足龍。編號PMU 24706(之前是PMU 234)包含九節關節連接的背椎和薦椎、兩根背肋、接近完整的骨盆、以及右後肢缺乏第五節蹠骨和多個趾骨[6]。
兩具標本都存放於瑞典烏普薩拉的烏普薩拉大學演化博物館,安裝好的骨架自1930年代開始展示。
1923年,師丹斯基缺乏足夠時間來完成正模標本的挖掘[4]。於是在1934年,楊鍾健和卞美年重返挖掘地並挖出四節後背椎、左肩胛和左肱骨[7]。這具標本由Wilson & Upchurch(2009)標註為「exemplar c」。他們認為這些骨骼來自與正模標本的同一個體。2007年的消息表示該標本可能不再屬於中國館藏[1]。
敘述

體型
盤足龍最早被敘述成大型的蜥腳類,成年體重約15至20噸及長15公尺。後來的研究將估計值縮小了。2016年葛瑞戈里·S·保羅估計體長11公尺及重3.5噸[8]。
解剖學特徵
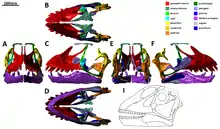

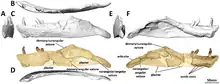
最早由衛曼所作的特徵檢驗已過時。檢驗是指針對特定物種(或類群)進行解剖學特徵的分析,特徵中可能包含獨有衍徵使物種與其他物種區別開來。
根據2009年Jeffrey A. Wilson及Paul Upchurch的研究,盤足龍可由以下獨有衍徵來鑑定[1]:
- 牙齒向前傾斜,以牙齒前端的琺瑯質邊緣、方向朝前方及上方、扶壁也更靠近上方為證。
- 樞椎(第二頸椎)神經棘末端有個空洞,裡面有三個更深的氣囊凹陷。
- 後樞椎(第三頸椎)有多樣發育的骨骺(epipophyses)以及前關節突(prezygapophyses)之下更細微的前骨骺(pre‐epipopophyses),在椎體和前關節突之間嶺脊的前部突起。
- 椎弓(neural arches)有骺的/前關節突的椎板(epipophyseal‐prezygapophyseal lamina),一道從骨骺延伸到前關節突間平行的嶺脊。以分開神經棘基部通常的凹陷來分隔開兩個囊狀區(pneumatocoels)。
- 頸椎的側凹縮成較小的小孔開口。
- 頸椎的神經棘在高度及長度上縮減。
- 第三頸椎的神經棘有一個橫向扁平化的前向突起。
- 前部頸椎在結節(tuberculum)與頭狀突(capitulum)有三個連接肋骨的刺為肋骨頭。
- 由於一個附生稚骨下橫突和兩個肋骨頭之間的一長段,頸肋牢固地位於椎骨下方。
- 後頸部和前背部的中薦前椎神經棘分裂或分叉,並且在頸部基部和前段背椎長有一個中位結節至少和幹骺(metapophyses)一樣大,分叉上的分叉形成三叉狀。
- 中段和後段背部parapophyseal/diapophyseal椎板排列成K字形。
- 薦前椎氣腔延伸至髂骨,而由氣囊貫穿。
分類

衛曼在1929年不確定盤足龍(當時叫作Helopus)的親緣關係而建立了獨立的盤足龍科(Helopodidae)[4]。楊將其修改成盤足龍亞科(Helopodinae),先是歸類在圓頂龍科(當時叫作Morosauridae),而後改至腕龍科。羅默在1956年建立了新的盤足龍亞科(Euhelopodinae)[5]。1990年,John Stanton McIntosh將盤足龍放置在圓頂龍科中[9]。
Wilson & Upchurch (2009)根據支序分析將盤足龍歸類於盤足龍科(Euhelopodidae),定義為一群起源於侏儸紀東亞地理隔絕情況下產生的特有種演化支。不確定盤足龍科是否為單系群[1]。傳統上常將盤足龍、峨嵋龍、馬門溪龍、蜀龍歸類於單系群盤足龍科的理論已不被近年的系統發生學研究所支持[10]。盤足龍具有類似於泰坦巨龍類的演化特徵,但比泰坦巨龍類原始,可能位於多孔椎龍類中的基礎位置。現在對盤足龍科的定義為:巨龍形類中,離盤足龍較近,而離內烏肯龍較遠的所有物種。[11]
以下的演化樹根據José L. Carballido, Oliver W. M. Rauhut, Diego Pol and Leonardo Salgado (2011)呈現盤足龍在大鼻龍類的大約位置:[10]

| 圓頂龍形類 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
以下盤足龍科演化樹則取自D'Emic(2012),呈現出盤足龍的親緣物種:[11]
| 盤足龍科 |
| |||||||||||||||||||||||||||
古生態學
出土與年代
盤足龍的模式標本發現於中國山東省的蒙陰組,標本出土於黃綠色砂岩和黃綠色粉砂岩。
蒙陰組的年代為貝里亞階-凡藍今階[12]。過去曾被認為是下白堊紀的巴列姆階或阿普第階,約1.29到1.13億年前。在1990年代曾被誤認為是侏羅紀晚期。
生活型態
衛曼的研究非常關注盤足龍的生活模式。雖然盤足龍的腳沒有其他蜥腳類寬,但衛曼描述的扁平足形象令人印象深刻,這是使具有非常重的柱狀腿能夠步態穩定的適應。他將這種扁平足聯想到當時流行的理論,即蜥腳類在水底行走,並以長頸作為通氣管,這可以解釋鼻孔的高位。他並使用馬和農民用的瑞典雪靴照片搭配說明來強調比較。然而衛曼承認此理論的爭議性,他已意識到在水中覓食柔軟多汁植物,其營養價值小,因此傳統觀點是不太可能的。[2]

至今水棲蜥腳類的觀點已完全推翻,牠們被視為典型的陸棲動物。某些蜥腳類似乎具有適應覓食底植的特徵,例如下垂的頭部、背部傾斜、頸部基部幾乎未朝上。有觀點認為所有蜥腳類頸部都需維持水平以防高血壓,且受到頸椎相互活動性所限制。根據Christian(2010)的研究以盤足龍作為蜥腳類抬高頸部向高處覓植的佐證。相對於後腿,前腿可能較長,如此可能是為了使頸部基部提高。他發現盤足龍頸部結構很適合傾斜。並估計頸部長4.6公尺,頸部加頭部的重量為210公斤。頸椎在40度至50度間時壓力最平均,並可像長頸鹿一樣維持筆直的姿勢。高血壓本來會消耗更多體力,但Christian計算出將頸部抬高五分鐘會比走路超過十五公尺來得更有效益。若可於五分鐘內吃掉的一株植物平均相距超過十五公尺,則往更高處尋找營養將更有效率。他還指出食物短缺時,有無抬高頸部的能力就成為爭取更多進食機會或只能挨餓的差別了。[13]
從盤足龍的飲食也可印證頸部高端姿勢。短頭顱、高下頜和寬湯匙形牙齒非常適合咬下針葉樹或種子蕨。從Poropat(2013)的研究得到印證:盤足龍食用高大而堅固的植被,並不同於梁龍超科或泰坦巨龍類的耙狀牙齒,佔有不同的生態棲位。[3]Salakka(2014)也證實這點,但他指出盤足龍的咬合力肯定比不上圓頂龍,且由於體型小,其絕對營養範圍也受限。頜骨只能進行簡單的上下咬合,幾乎沒有咀嚼能力。牙齒不只能夠咬斷、還能稍微割裂植物。[14]
參考資料
- Wilson, Jeffrey A.; Upchurch, Paul. . Journal of Systematic Palaeontology. 2009, 7 (2): 199–239. doi:10.1017/S1477201908002691.
- H. C. T'an. 1923. New research on the Mesozoic and early Tertiary geology in Shantung. Geological Survey of China Bulletin 5:95-135
- Poropat, Stephen F.; Kear, Benjamin P. . PLOS ONE. 2013-11-21, 8 (11): e79932. PMC 3836988
 . PMID 24278222. doi:10.1371/journal.pone.0079932.
. PMID 24278222. doi:10.1371/journal.pone.0079932. - C. Wiman. 1929. "Die Kreide-Dinosaurier aus Shantung" [The Cretaceous dinosaurs from Shantung]. Palaeontologia Sinica, Series C 6(1): 1-67
- A.S. Romer. 1956. Osteology of the Reptiles, University of Chicago Press 772 pp
- "Euhelopus." In: Dodson, Peter & Britt, Brooks & Carpenter, Kenneth & Forster, Catherine A. & Gillette, David D. & Norell, Mark A. & Olshevsky, George & Parrish, J. Michael & Weishampel, David B. The Age of Dinosaurs. Publications International, LTD. p. 70. ISBN 0-7853-0443-6.
- C.-C. Young. 1935. "Dinosaurian remains from Mengyin, Shantung". Bulletin of the Geological Society of China 14(4): 519-533
- Paul, G.S., 2016, The Princeton Field Guide to Dinosaurs. Second Edition, Princeton University Press p. 224
- McIntosh, J.S. 1990. "Sauropoda". Pp. 345-401 in: Weishampel, D.B., Dodson, P. & Osmólska, H. (eds) The Dinosauria. University of California Press, Berkeley
- José L. Carballido; Oliver W. M. Rauhut; Diego Pol & Leonardo Salgado. . Zoological Journal of the Linnean Society. 2011, 163 (2): 605–662. doi:10.1111/j.1096-3642.2011.00723.x.
- D'Emic, Michael D., (PDF), Zoological Journal of the Linnean Society, 2012, 166 (3): 624–671 [2020-10-25], doi:10.1111/j.1096-3642.2012.00853.x, (原始内容存档 (PDF)于2020-08-23)
- Xu, Jianqiang; Li, Zhong. . Journal of Asian Earth Sciences. 2015, 111: 284–301. doi:10.1016/j.jseaes.2015.07.008.
- Christian, A. . Biology Letters. 2010, 6: 823–825.
- Seela Salakka. (PDF). Uppsala universitet, Institutionen för geovetenskaper Examensarbete E, Paleobiologi. 2014 [2020-10-25]. (原始内容存档 (PDF)于2020-10-27).