歐羅巴龍屬
歐羅巴龍(屬名:Europasaurus,意為「歐羅巴蜥蜴」,或譯為歐洲龍)是一屬四足步行的植食性蜥腳類恐龍,屬於基礎大鼻龍類的腕龍科。生存於侏儸紀晚期啟莫里階,地質年代約1億5400萬年前,分布於當時的歐洲群島,相當於現今的德國北部。在小島的隔離環境中生活,造就了相當小的體型,只有6公尺長,成為島嶼侏儒化的典型實例。模式種兼唯一種豪氏歐羅巴龍(Europasaurus holgeri)於2006年由古生物學家奧克塔韋歐·馬諦烏斯等人命名、發表。[1]
| 欧罗巴龙属 | |
|---|---|
 | |
| 歐羅巴龍骨架模型 展示於瑞士阿塔恐龍博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | †蜥脚形亚目 Sauropodomorpha |
| 下目: | †蜥腳下目 Sauropoda |
| 科: | †腕龍科 Brachiosauridae |
| 属: | †欧罗巴龙属 Europasaurus Mateus et al. in Sander et al., 2006 |
| 模式種 | |
| †豪氏歐羅巴龍 Europasaurus holgeri Mateus et al. in Sander et al., 2006 | |
發現歷史

1998年,私人化石收藏家豪哲·魯克(Holger Lüdtke)在德國歐克、哈林爾霍德、哥廷爾霍德小鎮之間的朗根堡山一處採石場發現了單獨的蜥腳類牙齒。朗根堡白堊採石場已活躍了一個多世紀,透過爆破方式開採岩石,並主要加工製成肥料。採石場暴露出幾乎一整片連續面、厚達203公尺、屬於孫鐵層沉積岩的碳酸鹽礦床。[2]地質年代橫跨侏羅紀晚期早牛津階至晚啟莫里階,形成於水深不超過30公尺的淺海沉積環境。採石場裸露的岩層近乎呈垂直、略有倒轉,是由相鄰的下白堊紀哈茨山隆起所造成。這個採石場在地質學界名聲遠播,數十年來持續受到廣泛關注和研究,並吸引地質學系學生前來參觀。雖然盛產海相無脊椎動物化石,陸生動物化石卻相對罕見;這個蜥腳類牙齒的發現,成為來自德國北部侏儸紀首次發現的蜥腳類恐龍標本。[3]
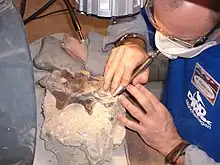
在更多化石材料(包括骨骼)被發現後,於1999年4月起,由當地私人化石收藏家協會發起含有骨骼岩層的挖掘活動。雖然採石場營運人員的配合度高,受到地層近乎垂直的走向、以及正在進行中的採石工作所限制,使得進入開鑿變得相當困難。蜥腳類化石材料無法直接從地層挖掘出來,而必須改以爆破方式,將岩石炸成碎片並從中取出。於是要考證骨骼材料的來源地變得較為困難,但後來的研究顯示,大概可以追溯回單一塊岩床-第83號岩床。[4]2000年7月20日至28日期間進行的挖掘工作,採集到約50公噸含有脊椎動物遺骸的岩石碎塊。[5]之後化石被運往鄰近漢諾威的戶外式私人恐龍博物館-慕尼黑恐龍公園(DFMMh)進行清修工作及典藏。由於骨骼保存狀態良好,固結劑僅能偶爾使用到,且骨骼很容易與周圍岩石分離,使得清修工作能夠以相對較快的速率完成。形狀較簡單的骨骼有時可以在不到一小時內就清修完畢,至於薦骨則耗時較長,約三個禮拜。截至2001年1月,已經有200塊單獨的骨頭清修完成。此時在面積為70×70公分的岩塊中發現的最密集的骨骼,包含約20塊。[4]到了2002年1月,在清修一塊更大的岩塊時發現了部分蜥腳類頭骨,亦成為歐洲發現過的首具蜥腳類頭骨。在將頭骨從石塊完全清出之前,先透過矽材質的印模來精準記錄下每塊骨骼的3D位置。[5]

2003年10月4日至5日夜間發生了縱火事件,燒毀了部分歐羅巴龍化石。該場火災毀壞了慕尼黑恐龍公園的實驗室與展廳,造成106塊骨頭遺失,約佔當時清修好骨頭的15%。此外火災也影響到多數尚未清修完成的石塊,消防水擊中高溫岩石時導致進一步碎裂。[6]毀壞的化石有DFMMh/FV 100標本,其中包含保存最完好的系列脊椎與唯一完整的骨盆。[7][6]

2006年正式發表敘述為蜥腳類的新屬新種,模式種兼唯一種是豪氏歐羅巴龍(Europasaurus holgeri)。屬名意為「來自歐洲的爬行類」,種名紀念化石最初發現者豪哲·魯克。[1]根據頭骨相對小型的尺寸,起初推測來自幼年個體。然而2006年的研究確認大多數標本都已成年,因此歐羅巴龍是島嶼侏儒化的產物。[3]此時個體骨骼數量增加至650塊,並包含數隻關節連接的個體;化石材料在60至80平方公尺的區域內發現。[2]從這些標本中指定出正模標本為DFMMh/FV 291,是一隻關節脫落但相關的個體骨骸,包含多塊頭骨碎片(前上頜骨、上頜骨、方軛骨)、部分腦殼、數塊下頜骨(齒骨、上隅骨、隅骨)、大量牙齒、頸椎、薦椎、屬於頸部和身軀的肋骨。根據重複部位的化石來判斷,至少還有其他10隻個體被歸入歐羅巴龍。[2]
2012年夏季再度展開大規模挖掘活動,目標為不僅僅從石塊,而是直接從岩層中挖出歐羅巴龍的骨骼。要進入到含有骨骼的地層區,須使用挖土機、有輪載土車來移除重達600公噸的岩石,並不斷從採石場底部進行抽水。挖掘工作到2013年春季及夏季持續進行著。此次活動的收穫包含新的魚類、龜類、鱷類遺骸,以及關於含骨層的寶貴資訊,然而找到額外的歐羅巴龍化石無法被定位。截至2014年,第83號岩床一共產出約1300具脊椎動物骨骼,其中大部分都屬於歐羅巴龍;估計還有3000塊額外骨骼尚待清修。根據頜骨數量,鑑定出至少20隻個體。[3]
描述
體型

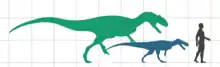
歐羅巴龍是相當小型的蜥腳類,成年個體身長估計僅有6.2公尺。這項估計是基於一個部分股骨,縮放至幾乎完整圓頂龍標本的比例。較年輕的個體也已得知,從3.7公尺長到最幼年的1.75公尺長都有。[1][2]同樣侏儒化的泰坦巨龍類馬扎爾龍有著相近的體型,體重為900公斤,而歐羅巴龍體重約800公斤。另一種非侏儒化的最小型泰坦巨龍類內烏肯龍的紀錄依然超越前兩者,身長9公尺及體重3.2公噸。[8]
鑑定特徵
根據初始敘述論文,歐羅巴龍除了作為一種體型非常小的新蜥腳類,也有足夠的數相鑑定特徵可以和近緣物種區別開來:前上頜骨鼻突前彎且上突(現在知道是人工復原[9]);頸椎椎體上面有個孔洞;肩胛骨後面有個明顯突起;距骨寬度為長度的兩倍。[1]
與圓頂龍進行比較:歐羅巴龍具有不同的眶後骨形態,其中後緣沒那麼短;鼻骨和額骨之間有個很短的接觸面;頂骨形狀為矩形;骨盆以前的脊椎神經棘沒有分叉。與當時稱為腕龍(現在是長頸巨龍)進行比較:歐羅巴龍有個較短的吻部;方軛骨與鱗狀骨間有接觸;肱骨近端與遠端面平坦且對稱。與葡萄牙巨龍進行比較:兩者髂骨和距骨形狀不同。與多里亞巨龍進行比較:後者三角肌嵴沒那麼突出,延伸到肱骨程度較少。[1]
頭骨

幾乎整個歐羅巴龍頭骨外部部位都保存下來了,除了前額骨沒有。某些部位骨頭僅有非常破碎、資訊量不足的化石,如淚骨。已知有八個前上頜骨,吻部形狀大致呈矩形,如同在圓頂龍觀察到的情況。2006年鑑定出前上頜骨的前突,[1]2014年重新確認是人工復原出來的,[9]較少程度上類似於在圓頂龍、盤足龍觀察到的解剖結構。前上頜骨與鼻骨接觸的背突起緣於後背突,然後在鼻下孔處變得筆直且垂直,直到最後接觸鼻骨。這樣微弱的過程也可見於圓頂龍和盤足龍,並在阿比杜斯龍、長頸巨龍及可能的腕龍頭骨中更為明顯。後面幾種的吻部較長,從第一顆牙齒到前上頜骨鼻突間的距離更大。另外歐羅巴龍的眼眶、鼻孔尺寸與基礎圓頂龍形類(腕龍科、圓頂龍、盤足龍、馬拉威龍)較接近,而較進階的泰坦巨龍類(掠食龍、塔普亞龍、納摩蓋吐龍)鼻孔尺寸大幅縮小。[9]

在保存良好的歐羅巴龍標本DFMMh/FV 291.17中有個單獨的上頜骨。其骨體長並有兩個突起:一個鼻突和一個後突。只有一道發育很弱的淚突,如同大部分蜥腳類,除了掠食龍。鼻突加長並包覆眶前孔的前腹緣,這個突起從齒列水平方向延伸約120度角。鼻突基部亦構成了淚突的骨體,而眶前孔成為彼此的分隔點,形狀類似圓頂龍、盤足龍、阿比杜斯龍、長頸巨龍,但比例上高了約1/2。眶前孔前面或下面的前眶前孔幾乎消失,類似圓頂龍及盤足龍,而不像在梁龍、塔普亞龍的發育良好。歐羅巴龍上頜骨總共有12至13顆牙齒,比更基礎的物種(約巴龍16顆、亞特拉斯龍14至25顆)還少,但落在腕龍科的變化值範圍內(腕龍15顆、阿比杜斯龍10顆)。所有保存下來未磨耗的牙齒近中緣都顯示出多達四個小齒突。少數後部齒冠略微扭轉(~15度),但遠小於腕龍科(30至45度)。[9]

在保存下來多個歐羅巴龍鼻骨中,很少是完整或未變形的。鼻骨後部被額骨覆蓋,往側邊約略與前額骨關節相連。不像長頸巨龍的額骨,歐羅巴龍的額骨水平向前突,構成眶前孔上方頭蓋骨的一小部分。已知的歐羅巴龍額骨有四個,其中三個來自左邊,一個來自右邊。因為關節脫落,額骨在發育階段可能從未癒合,不像圓頂龍的情況。額骨構成頭蓋骨的一部分,與其他骨頭如鼻骨、頂骨、前額骨、眶後骨關節相連,其前後長度長於寬度,在真蜥腳類是一項獨特的特徵。如同梁龍超科(阿馬加龍、叉龍、梁龍)以及圓頂龍,額骨被排除於額頂孔(或稱頂骨孔,因為額骨排除在外)之外。額骨也被排除於上顳孔(supratemporal fenestra)邊緣之外,在比蜀龍進階的蜥腳類中是普遍的特徵;且額骨在眼眶只有很小、無裝飾性的參與構成。已知有很多個歐羅巴龍頂骨,形狀呈矩形,寬度大於長度。從頭骨後方視角頂骨也很寬,位置略高於枕骨大孔。頂骨參與構成後顳孔(頭骨最後面的孔洞)邊緣大約一半,其他區域則與鱗狀骨和一些腦殼骨頭靠攏。頂骨亦構成上顳孔部分邊緣,其寬度大於長度,如同長頸巨龍、圓頂龍、棘刺龍。除了前述幾個孔洞,頂骨本身也具有一個「後頂孔」,這在除了叉龍科之外是相當罕見的特徵。歐羅巴龍具有三條放射狀的眶後骨,是從更基礎物種中的後額骨和眶後骨癒合演變而來。在前側和腹側突起之間眶後骨構成了眼眶邊緣,並在腹側和後側突起之間眶後骨分隔了顳顬孔。[9]

已知有很多個歐羅巴龍顴骨,比起其他大鼻龍類,反而與基礎蜥腳形亞目更為類似。顴骨構成眼眶、下顳孔、顱底緣的一部分,但未達到眶前孔的位置。顴骨後突非常纖弱及狹窄,在與方軛骨關節相接處顯示出骨質凹痕。顴骨骨體後方有兩道突出,以75度角分叉,構成下顳孔底部與前緣。如同里奧哈龍、大椎龍這兩種非蜥腳下目的蜥腳形亞目,顴骨構成眼眶緣很大一部分,從後面到前面底部角落。此特徵在泰坦巨龍類的胚胎有觀察到,但不見於成體。方軛骨是條細長的骨頭,有兩個突出的枝,一條在前一條在背。如同其他蜥腳類,前突長於背突,但歐羅巴龍兩枝長度較接近。水平突與齒列平行,類似圓頂龍,但不像長頸巨龍和阿比杜斯龍。骨頭前枝有個突出的腹緣,可能是腕龍科的共有衍徵,但也存在於某些圓頂龍個體。兩根方軛骨突以近乎90度直角分叉,背突因沿方骨形狀而呈現彎曲狀。側視歐羅巴龍的鱗狀骨看到與圓頂龍近似的問號形狀。鱗狀骨與其他許多塊頭骨關節相連,包含頭蓋骨、腹側頭骨及腦殼。如同眶後骨,鱗狀骨呈三條放射狀,具有腹突、前突、內突。[9]
有十三個歐羅巴龍的腭部材料保存下來,包含方骨、翼骨、齶骨。方骨與腭部、腦殼、外部頭骨關節相連,形狀與長頸巨龍、圓頂龍相似,都有發育良好的關節面。方骨長邊大部分為單個軸,內側有個翼狀。翼骨是蜥腳類腭部最大的骨頭,具有三條放射狀,如同眶後骨和鱗狀骨。一條前突接觸翼骨相對位置、一條側翼接觸齶骨、一條後翼支撐方骨和基翼骨(腭部和腦殼相連的骨頭)。齶骨是一塊小腭骨,將中央腭骨(翼骨、齶骨)與上頜骨關節連接在一起。齶骨呈L形,前突接觸上頜骨,背突與翼骨接壤。[9]
脊椎

頸椎是歐羅巴龍脊柱中保存最良好及最多的部分。但不是整條頸部都已知,因此頸椎數目可能介於圓頂龍(12節)與掠食龍(17節)之間。此外來自不同年齡層個體的數個頸椎,使能夠瞭解到椎體長度和內部結構會隨發育過程而改變。成體頸椎椎體加長且為雙後凹(opisthocoelous),即前端呈球狀、後端凹陷;椎體後端頂部有個凹槽。這在歐羅巴龍被視為典型特徵,但也可在盤足龍和長頸巨龍身上觀察到。歐羅巴龍椎體側面有個挖入的凹陷,通往椎體內部的氣腔。不像長頸巨龍和其他腕龍科,歐羅巴龍這邊的開口不具有狹窄的椎板(laminae)作分隔。歐羅巴龍脊椎上部的椎板特徵與基礎大鼻龍類和腕龍科有相同之處。與前中部頸椎不同的是,後部頸椎加長幅度較小,比例上較高,如同其他大鼻龍類,關節面位置產生明顯的變化。[7]


前部背椎與頸椎一樣呈現強烈的雙後凹,並可根據下椎弓凹(hypantrum)的缺乏與椎體側突(parapophysis)的低位置來判斷在脊柱中的定位。內部結構如同圓頂龍、腕龍、加爾瓦龍呈現開放式、分室(camerate),但氣腔沒有延伸到背椎中部和後部,不同於前述幾者。歐羅巴龍的前椎板形式與其他基礎大鼻龍類相似,但不同於更基礎(如馬門溪龍、簡棘龍)及更進階(如長頸巨龍)的物種。中部背椎有個氣腔向上延伸至神經弓,類似巨腳龍、鯨龍、特維爾切龍、圓頂龍。此開孔的腹緣呈菱形且輪廓分明。後部背椎的側凹位移至椎體更高處,此變化也可在其他基礎大鼻龍類觀察到。這些側凹前寬後窄形成銳角。歐羅巴龍的神經棘垂直豎立,是個在腕龍或其他更進階大鼻龍類中所沒有的基礎特徵。[7]
DFMMh/FV 100保存了唯一已知完整的薦椎系列,但已於2003年的大火中燒毀。所有五節薦椎(較基礎新蜥腳類的典型數目)都與薦骨癒合在一起。第三和第四節薦椎等同原本的薦骨,是所有恐龍的共通特徵。第二薦椎是從基礎蜥腳形亞目開始加入的祖先型薦骨,牠們都有三節薦骨,除了板龍。第五薦椎癒合於原生對列之後,稱為尾薦,從尾部移至骨盆,存在於利奧尼拉龍周邊物種。第一薦椎與髂骨相連,但不與其他薦椎接觸,是為背薦,使所有新蜥腳類都可觀察到薦椎數目達到五個。背薦的癒合程度可用來確認蜥腳類薦椎數目的演化過程:最初原生的一對加上一個背椎(三個),然後加入一個尾椎(變四個),最後多加一個背椎(變五個)。[7]
皮膚

在大鼻龍類中,已知只有黑斯廷龍、特維爾切龍、薩爾塔龍保存了化石化的皮膚印痕。特維爾切龍和黑斯塔龍可能與歐羅巴龍近緣,且所有蜥腳類皮膚特徵都很類似。黑斯廷龍是第一個已知保存皮膚印痕的恐龍,在肘關節周圍的部分手臂保存了表皮。[10]真皮印痕在特維爾切龍材料中分布更廣泛,所知來自前肢、肩胛、身軀,沒有代表裝甲的骨板或結節,但有數種類型的鱗片。特維爾切龍的皮膚種類整體上更類似梁龍科和黑斯廷龍,而比較不像奧卡馬韋渥發現的泰坦巨龍類胚胎。[11]由於這些基礎大鼻龍類保存下來的結節形狀及關節與其他也有保存皮膚的物種相似,包括秀麗雷龍、梁龍科未定物種,因此推測這種皮膚結構可能普遍存在於所有新蜥腳類中。[10]
分類
分類為基礎大鼻龍類
起初命名時歐羅巴龍被認為屬於大鼻龍類,但沒有落在腕龍科或巨龍形類的範圍內。這代表著歐羅巴龍的侏儒化是演化的產物,而非該類群的特徵。[1]包含進歐羅巴龍的系統發生學分析採納了三份研究數據:威爾森(2002)[12]、阿普撤奇(1998)[13]、阿普撤奇等人(2004)[14]。所有分析都得出相似的結果,其中歐羅巴龍比原頂龍還要衍化,但被排除於腕龍科和巨龍形類之外。山德等人(2006)偏好的結果如下左邊演化樹所示:[2]
|
|
在卡巴利多和山德(2013)描述歐羅巴龍脊椎的論文中進行了另一項系統發生學分析,結果如上右邊演化樹所示。支序範圍擴張到包含進更多蜥腳類物種,如巧龍、雪松龍、塔普亞龍。根據泰勒(2009),腕龍屬也被拆分為真正的腕龍(高胸腕龍)和長頸巨龍(布氏腕龍)。[15]依照這個更廣泛的更新版本,歐羅巴龍被發現落在與之前相似的位置,即離具龍形類較接近,而離圓頂龍較遠的基礎圓頂龍形類。但盤足龍、特維爾切龍、塔斯塔維斯龍和加爾瓦龍都位於歐羅巴龍和腕龍科之間的位置。[7]
分類為腕龍科
狄米克(2012)的巨龍形類系統發生學分析認為歐羅巴龍屬於腕龍科,而不是比最早期腕龍科還基礎的物種。研究將腕龍科定義修正為目前所知最擬真的版本,雖然仍有數種潛在腕龍科後來被確認屬於更進階的多孔椎龍類(帕拉克西龍、波塞東龍、橋灣龍)。但狄米克也只暫時將歐羅巴龍列在確定的腕龍科。就算系統發生分析對歐羅巴龍位置提供了有力支持,牠作為少數有發現頭骨的大鼻龍類,卻缺乏數個腕龍科應有的共有衍徵,例如尾椎方面。左下為狄米克(2012)的系統發生演化樹:[16]
|
|

後續的巨龍形類分析認同狄米克(2012)的歐羅巴龍分類。牠與腕龍、法國的溝椎龍(現在命名成蝰神龍)一同於腕龍科最基礎的位置構成多分支,下一階較進階的分支是葡萄牙巨龍,而長頸巨龍、阿比杜斯龍、雪松龍、毒癮龍構成腕龍科一個最進階的演化支。歐羅巴龍的扭曲型牙齒被發現是腕龍科的獨有特徵之一,這意味著可能透過這項可靠的依據而將一些只有單獨牙齒的蜥腳類物種歸類進腕龍科。[17]
狄米克等人(2016)以2012年的分析作為基礎,進行了進一步的系統發育分析,得出歐羅巴龍在腕龍科中與之前幾乎一樣的位置,但索諾拉龍與長頸巨龍構成一個演化支,而葡萄牙巨龍則被移到與阿比杜斯龍、雪松龍一同構成的多分支。支序圖其餘部分與2012年相同,然而腕龍被放進更進階的腕龍科多分支中。[18]曼尼翁等人(2017)另一版本的系統發育分析得出與狄米克(2012)和狄米克等人(2016)相似的結果。歐羅巴龍是最基礎的腕龍科,法國溝椎龍/蝰神龍是第二基礎的腕龍科。腕龍被排除於其他所有腕龍科(長頸巨龍、阿比杜斯龍、索諾拉龍、雪松龍、毒癮龍)構成的多分支之外。[19]羅約托雷斯等人(2017)得出腕龍科中更複雜的親緣關係。除了歐羅巴龍作為最基礎的物種,腕龍科底下還區分成兩個次演化支,一個包含長頸巨龍、索諾拉龍、葡萄牙巨龍,另一個包含其餘所有腕龍科以及塔斯塔維斯龍。根據對腕龍科的定義,第二個演化支被稱為勞亞龍形類。腕龍與這兩個次演化支一同形成多分支。羅約托雷斯等人(2017)的演化樹如右上方所示。[20]
古生物學
發育

根據對數個歐羅巴龍標本的組織學分析研究顯示,牠並不是其他已知物種(如圓頂龍)的幼年體,而是一個獨特的侏儒化物種。透過這方面的分析顯示出,最年輕、體型最小(僅長1.75公尺)的DFMMh/FV 009標本缺乏成長中的跡象,如發育痕跡或年輪組織。這些組織是表明快速生長的證據,因此該標本可能是個年輕的幼體。稍大一點長2公尺的DFMMh/FV 291.9標本顯示出大量年輪組織,但缺乏發育痕跡,因此可能也是個幼體。第三小的DFMMh/FV 001標本身長3.5公尺,存在發育跡象(尤其是環狀),推測可能是個亞成體。再大一些的DFMMh/FV 495標本身長3.7公尺,呈現出成熟的骨單位以及環狀痕跡。受檢驗第二大型的DFMMh/FV 153標本身長4.2公尺,同樣顯示出發育痕跡,但頻率更高。單一個部分股骨標本DFMMh/FV 415是已知最大型的歐羅巴龍個體,身長6.2公尺;不像其他標本,這個標本呈現出成長停滯線,代表牠死時已經達到完全成長的體型。骨骼內部亦呈現部分層狀痕跡,代表最近已停止發育。[1][2]
這些生長因子的組合證明,在增長率大幅降低的情況下,歐羅巴龍發育出如此小的體型,相較於其他大型物種(如圓頂龍)的體型增長速率更慢。此種趨緩的增長率與蜥腳類和獸腳類普遍趨勢相違背,正常情況的後者會隨增長率的提高而達到更大的體型。[1]某些歐羅巴龍的近親,如腕龍、波塞東龍是已知最巨型的恐龍代表。馬曼等人(2014)認為歐羅巴龍的小體型和減緩增長率是種稚態的展現,意即成體仍保留了幼體的特徵,例如體型。[9]
侏儒化

有一種理論認為,當歐羅巴龍的祖先遷徙至當時存在的島嶼後,體型便發生了迅速縮小,因為相當於現在德國北部附近當時存在的島嶼,面積最大不超過20萬平方公里,環境負載力可能無法支撐大型蜥腳類群落。另一種說法是,大鼻龍類的體型也可能隨著遷徙至越來越小的島嶼而逐漸變小,直到最後達到歐羅巴龍的體型。[1]過去對於島嶼侏儒化恐龍的研究主要關注於羅馬尼亞馬斯垂克階的哈提格島,是侏儒泰坦巨龍類馬扎爾龍和侏儒鴨嘴龍類沼澤龍的棲息地。[1][21][22]沼澤龍的了解來自一隻小型成體,而馬扎爾龍的標本雖然非常小型,卻也是來自成體甚至是年老個體。[22][8]達契亞馬扎爾龍成體體型接近歐羅巴龍,但後者最大個體的股骨長於前者,而匈牙利馬扎爾龍則明顯大於前兩者。[8]歐羅巴龍的侏儒化代表著在進階蜥腳形亞目中唯一已知的快速體重顯著下降,因為其他物種的普遍趨勢都是整體體型的增長。[23]
古生態學
環境與共棲物種

德國的朗根堡點,年代橫跨早牛津階到晚啟莫里階,展現出侏羅紀晚期島嶼生態系統的動植物多樣性。[24]啟莫里階時的當地本來是一片海洋,為於萊尼施(Rhenisch)、波西米亞(Bohemian)、倫敦-布拉班特(London-Brabant)地塊之間。這不代表物種都是海洋生物,動植物可能從距離很近的周圍島嶼漂過來沉積。[25]沉積物顯示偶爾會有淡水或半鹹水流入,保存下來的化石也證明這點。採石場的石灰岩層產出大量海洋雙殼綱、棘皮動物、微生物化石,但發現的很多動植物則是陸生的。[26]
雖然朗根堡保存了很多海洋生物,牠們卻不太可能常態與歐羅巴龍共存。至少有三個以上的烏龜物種,包含近龜、地中海龜和至多兩種未命名。[24][27][28]輻鰭魚相當豐富,如鱗齒魚、Macromesodon、Proscinetes、Coelodus、Macrosemius、Notagogus、Histionotus、Ionoscopus、Ainia、Caturus、蜥目魚、巨鱵嘴魚、Thrissops。[29]還有至少五種形態獨特的弓鮫目、板鰓亞綱的Palaeoscyllium、古棘鯊、星齒魟。[30]兩種海生鱷形類鬥蜥鱷和狹蜥鱷,可能以魚類和海龜為食。亦有發現兩棲型的稜角鱗鱷。[31]

陸地方面的化石中,毬果和樹枝可被鑑定為南洋杉科的短葉杉屬。沉積的動物種類繁多,包括大量歐羅巴龍個體骨骼。朗根堡採石場發現了至少450塊歐羅巴龍骨骼,其中1/3帶有齒痕。[24]這些齒痕的尺寸及形狀與魚類、鱷類、其他食腐動物的牙齒非常匹配,但沒有一個確認是屬於獸腳類的。[24][32]大量個體的存在表明,一群歐羅巴龍在穿越潮汐帶時被淹死。[24]雖然目前發現主要的大型動物殘骸都是歐羅巴龍,也有一些來自梁龍類、劍龍類、許多種獸腳類的殘骸。三節梁龍類的頸椎有保存下來,從尺寸來判斷很可能也是個侏儒化物種。劍龍類和許多種獸腳類僅有牙齒保存,除此之外還有少數骨骼可能屬於角鼻龍科。[24]單獨的牙齒顯示出當地存在著至少四種不同的獸腳類,包括斑龍科蠻龍未命名種、另一種斑龍科、異特龍科和角鼻龍類未定種,甚至還有已知最古老的伶盜龍亞科牙齒。[33][34]

除了恐龍以外,朗根堡採石場還保存許多小體型的陸生脊椎動物,如翼龍類準噶爾翼龍科保存良好的立體骨骼、鳥掌翼龍類和梳頜翼龍科的單獨遺骸;一種Paramacellodidae科蜥蜴;一種野獸鱷近親的部分骨骼及頭骨,現在命名成諾曲奇鱷;[24][25][27]磔齒獸科化石亦有保存,還有一種柱齒獸類、一種始俊獸科、一種多瘤齒獸目。[24][35]
延伸閱讀
- Hall, B.K.; Hallgrímsson, B. . Jones & Bartlett Publishers. 2011: 446 [2021-08-30]. ISBN 978-1-4496-4722-3. (原始内容存档于2022-04-16).
- Brusatte, S.L. . John Wiley & Sons. 2012: 210 [2021-08-30]. ISBN 978-0-470-65658-7. (原始内容存档于2021-10-04).
- Bolecsek, C.; Wings, O. . 2013: 46–47. ISBN 978-84-695-8567-2.
|journal=被忽略 (帮助) - Organ, C.L.; Brusatte, S.L.; Stein, K. . Proceedings of the Royal Society of London B: Biological Sciences. 2009, 276 (1677): 4303–4308. ISSN 0962-8452. PMC 2817110
 . PMID 19793755. doi:10.1098/rspb.2009.1343.
. PMID 19793755. doi:10.1098/rspb.2009.1343. - Filippini, F.S.; Otero, A.; Gasparini, Z. . Alcheringa. 2016, 41: 69–78. ISSN 0311-5518. S2CID 131785420. doi:10.1080/03115518.2016.1180806.
參考來源
- Sander, P.M.; Mateus, O.V.; Laven, T.; Knötschke, N. . Nature. 2006, 441 (7094): 739–741. Bibcode:2006Natur.441..739M. PMID 16760975. S2CID 4361820. doi:10.1038/nature04633.
- Sander, P.M.; Mateus, O.V.; Laven, T.; Knötschke, N. . Nature. 2006, 441 (7094): 739–741. Bibcode:2006Natur.441..739M. PMID 16760975. S2CID 4361820. doi:10.1038/nature04633.
- Wings, Oliver. . Fossilien. 2014, 2: 44–50 (德语).
- Knötschke, Nils. . Der Präparator (Hannover). 2001, 47 (1): 7–13 (德语).
- Knötschke, Nils; Rössler, Dennis. . Der Präparator (Hannover). 2002, 48 (1): 21–30 (德语).
- Wings, O.; Knötschke, N. . 74th Annual Meeting of the Society of Vertebrate Paleontology: Program and Abstracts.
- Carballido, J.L.; Sander, M.P. (PDF). Journal of Systematic Palaeontology. 2013, 12 (3): 335–387 [2021-08-30]. ISSN 1477-2019. S2CID 85087382. doi:10.1080/14772019.2013.764935. (原始内容 (PDF)存档于2017-10-19).
- Stein, K.; Csiki, Z.; Curry-Rogers, K.; Weishampel, D.B.; Redelstorff, R.; Carballido, J.L.; Sander, P.M. . Proceedings of the National Academy of Sciences. 2010, 107 (20): 9258–9263. Bibcode:2010PNAS..107.9258S. ISSN 0027-8424. PMC 2889090 . PMID 20435913. doi:10.1073/pnas.1000781107 .
- Marpmann, J.S.; Carballido, J.L.; Sander, P.M.; Knötschke, N. . Journal of Systematic Palaeontology. 2014, 13 (3): 1–43. S2CID 84076044. doi:10.1080/14772019.2013.875074.
- Upchurch, P.; Mannion, P.D.; Taylor, M.P. . PLOS ONE. 2015, 10 (6): e0125819. Bibcode:2015PLoSO..1025819U. ISSN 1932-6203. PMC 4454574 . PMID 26039587. doi:10.1371/journal.pone.0125819 .
- Giménez, O. del V. (PDF). Revista del Museo Argentino de Ciencias Naturales. 2007, 9 (2): 119–124 [2017-10-25]. doi:10.22179/revmacn.9.303. (原始内容 (PDF)存档于2012-10-07).
- Wilson, J.A. (PDF). Zoological Journal of the Linnean Society. 2002, 136 (2): 217–276 [2021-08-31]. doi:10.1046/j.1096-3642.2002.00029.x. (原始内容 (PDF)存档于2022-02-09).
- Upchurch, P.M. . Zoological Journal of the Linnean Society. 1998, 124: 43–103. doi:10.1111/j.1096-3642.1998.tb00569.x.
- Upchurch, P.M.; Barrett, P.M.; Dodson, P. . Weishampel, D.B.; Dodson, P.; Osmólska, H. (编). 2. Berkeley: University of California Press. 2004: 259–322. ISBN 978-0-520-94143-4.
- Taylor, M.P. . Journal of Vertebrate Paleontology. 2009, 29 (3): 787–806. S2CID 15220647. doi:10.1671/039.029.0309.
- D'Emic, M.D. (PDF). Zoological Journal of the Linnean Society. 2012, 166 (3): 624–671 [2021-08-31]. doi:10.1111/j.1096-3642.2012.00853.x. (原始内容 (PDF)存档于2020-08-23).
- Mannion, P.D.; Upchurch, P.; Barnes, R.N.; Mateus, O. . Zoological Journal of the Linnean Society. 2013, 168 (1): 98–206. ISSN 0024-4082. doi:10.1111/zoj.12029.

- D’Emic, M.D.; Foreman, B.Z.; Jud, N.A. . Journal of Paleontology. 2016, 90 (1): 102–132. ISSN 0022-3360. S2CID 133054430. doi:10.1017/jpa.2015.67.
- Mannion, P.D.; Allain, R.; Moine, O. . PeerJ. 2017, 5: e3217. ISSN 2167-8359. PMC 5417094 . PMID 28480136. doi:10.7717/peerj.3217.
- Royo-Torres, R.; Fuentes, C.; Meijide, M.; Meijide-Fuentes, F.; Meijide-Fuentes, M. . Cretaceous Research. 2017, 80: 38–55. doi:10.1016/j.cretres.2017.08.012.
- Nopcsa, F. . Ver. Zool. Bot. Ges. Wien. 1914, 54: 12–14.
- Weishampel, D.B.; Norman, D.B.; Grigorescu, D. . Palaeontology. 1993, 36: 361–385.
- Benson, R.B.J.; Campione, N.E.; Carrano, M.T.; Mannion, P.D.; Sullivan, C.; Upchurch, P.; Evans, D.C. . PLOS Biology. 2014, 12 (5): e1001853. PMC 4011683 . PMID 24802911. doi:10.1371/journal.pbio.1001853.
- Wings, O. . Abstracts of the 12th Symposium on Mesozoic Terrestrial Ecosystems. 2015: 99–100.
- Schwarz, D.; Raddatz, M.; Wings, O. . PLOS ONE. 2017, 12 (2): e0160617. Bibcode:2017PLoSO..1260617S. PMC 5310792 . PMID 28199316. doi:10.1371/journal.pone.0160617 .
- Wings, O. . Journal of Vertebrate Paleontology, Program and Abstracts. 73rd Annual Meeting of the Society of Vertebrate Paleontology. 2013, 2013: 239.
- Wings, O.; Sander, M.P. . 10th Annual Meeting of the European Association of Vertebrate Paleontologists. 2012, 20: 281–284.
- Marinheiro, J.; Mateus, O. (PDF). 71st Annual Meeting of the Society of Vertebrate Paleontology. Abstracts of the 71st Annual Meeting of the Society of Vertebrate Paleontology. 2011: 151 [2021-08-31]. (原始内容 (PDF)存档于2021-10-04).
- Mudroch, A.; Thies, D. . Geologica et Palaeontologica. 1996, 30: 239–265 (德语).
- Thies, D. . Journal of Vertebrate Paleontology. 1995, 15 (3): 463–481. doi:10.1080/02724634.1995.10011242.
- Karl, H.-V.; Gröning, E.; Brauckmann, C.; Schwarz, D.; Knötschke, N. . Clausthaler Geowissenschaften. 2006, 5: 59–77.
- Slodownik, M.; Wings, O. . 13th Annual Meeting of the European Association of Vertebrate Palaeontologists Opole, Poland, Abstracts. 2015: 141.
- Gerke, O.; Wings, O. . PLOS ONE. 2016, 11 (7): e0158334. Bibcode:2016PLoSO..1158334G. PMC 4934775 . PMID 27383054. doi:10.1371/journal.pone.0158334 .
- Lubbe, T. van Der; Richter, U.; Knötschke, N. (PDF). Acta Palaeontologica Polonica. 2009, 54 (3): 401–408 [2021-08-31]. S2CID 38105783. doi:10.4202/app.2008.0007. (原始内容 (PDF)存档于2021-04-14).
- Martin, T.; Schultz, J.A.; Schwermann, A.H.; Wings, O. (PDF). Palaeontologia Polonica. 2016, 67: 171–179 [2021-08-31]. doi:10.4202/pp.2016.67_171 (不活跃 31 May 2021). (原始内容 (PDF)存档于2016-08-10).
- Lallensack, J.N.; Sander, M.P.; Knötschke, N.; Wings, O. (PDF). Palaeontologia Electronica. 2015, 18 (2): 1–34 [2021-08-31]. (原始内容 (PDF)存档于2022-02-20).