重爪龍屬
重爪龍屬(屬名:,發音:/ˌbæriˈɒnɪks/,意為「沉重的爪」,又名堅爪龍)為棘龍科的獸腳類恐龍,大約生存於1億3000萬至1億2500萬年前的早白堊世巴列姆階。首具化石於1983年發現於英格蘭薩里郡的威爾德黏土組,並成為模式種沃氏重爪龍(Baryonyx walkeri)的正模標本,由古生物學家亞倫·查理格和安琪拉·米爾納於1986年敘述、命名。屬名來自手部第一指非常巨大的爪子;種名紀念化石發現者:業餘化石收藏家威廉·沃克(William J. Walker)。重爪龍的正模標本是英國所發現最完整的獸腳類骨骼之一,並且一直是已知棘龍科中標本最完整的物種,牠的發現也引起了大眾媒體的關注。後續在英國其他地方以及伊比利半島發現的標本也被歸入本屬。
| 重爪龙属 化石时期:白堊紀早期, | |
|---|---|
 | |
| 重爪龍的重建骨架模型,展示於東京國立科學博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 獸腳亞目 Theropoda |
| 科: | †棘龍科 Spinosauridae |
| 亚科: | †重爪龙亚科 Baryonychinae |
| 属: | †重爪龙属 Baryonyx Charig & Milner, 1986 |
| 模式種 | |
| †沃氏重爪龍 Baryonyx walkeri Charig & Milner, 1986 | |
正模標本估計身長介於7.5至10公尺之間、體重約1.2至1.7噸,可能是個未完全成熟的個體。重爪龍擁有長、低矮而狹窄的吻部,經常被拿來與現代鱷魚相互比較。吻部尖端呈花座狀擴張至側面。其後的上頜有個凹口,與對應地區突起的下頜互相契合。鼻骨頂部有個三角形的冠。重爪龍有著大量帶細鋸齒邊緣、圓錐狀的牙齒,其中最大顆的牙齒位於前端。頸部呈S形,背部脊椎的神經棘由前向後逐漸變高。一根加長的神經棘代表延背部中線可能有駝峰或脊狀的結構。前肢強壯發達,屬名由來的第一指爪測量長約31公分。
在當初發現時,重爪龍的親緣關係充滿著謎,現在已確認屬於棘龍科的一員。一些研究人員認為刀齒鱷龍是本屬較早的異名;而泰內雷似鱷龍也屬於同一物種;後續的研究則繼續將牠們列為獨立的屬。重爪龍是第一個確認為食魚性的獸腳類恐龍,根據正模標本胃部發現的魚鱗得到證實。牠可能也是能獵捕較大型的獵物的活躍型掠食者、以及食腐動物,因為在化石體內還發現了少年禽龍類的殘骸。推測牠生前會以前肢和大爪來捕捉和處理獵物。重爪龍可能擁有半水生的生活型態,並與其他獸腳類、鳥腳類、蜥腳類恐龍、以及翼龍類、鱷魚、烏龜、魚類共享著河相棲息地。
發現歷史

1983年1月,英國水電工兼業餘化石收藏家威廉·沃克(William J. Walker)在英格蘭薩里郡奧克利附近屬於威爾德黏土組的泥坑煙囪坑進行探勘。他在一塊岩石中發現一根巨大的爪子,但已經碎裂,帶回家將其修復黏合後,才意識到爪子的尖端已經遺失。幾個星期後,沃克回到泥坑中的同一地點,經歷幾小時的搜索,找到了遺失的部分。他還發現趾骨和部分的肋骨。沃克的女婿將爪子帶到倫敦自然史博物館,在那邊由英國古生物學家艾倫·查理格和安琪拉·米爾納進行檢驗,並鑑定為屬於獸腳類恐龍。[1][2]古生物學家們於2月在同地點發現了更多骨骼碎片,但礙於泥坑內的天氣狀況,直到5、6月才採集全部的骨骼。[3][2]一個由8位博物館員工和幾名志工組成的團隊,在三週的時間內,在54個岩塊中挖出了重達2噸的岩基質。沃克將爪子捐贈給博物館,而擁有泥坑所有權的奧克利磚頭公司(Ockley Brick Company)捐贈了其餘骨骼並提供設備。[3][4][2]這個地區已經被探勘過超過200年的歷史,但過去從未發現過類似的遺骸。[5][2]
大部分採集到的骨骼都包裹在粉砂岩塊中,岩塊周圍由細砂和粉砂包覆,其餘部分則埋沒於泥土中。骨骼關節分離,並散布於5×2公尺的範圍內,但大多數都不遠離原本的位置。推土機打亂了骨骼的位置,有些骨骼在採集前就被機械設備破壞掉。[3][1][6]因為粉砂岩基質十分堅硬以及存在菱鐵礦,使清修標本變得非常艱難;曾嘗試用強酸清理,但大部分基質還是以機械方式去除。將所有骨骼從岩塊中清出,前後共耗時六年不間斷的清修,到最後階段的時候,還需在顯微鏡下使用牙科工具和氣錘來清修。標本約佔整個身體骨架的65%,包含部分頭骨:前上頜骨、左上頜骨、成對的鼻骨、左淚骨、左前額骨、左眶後骨、腦殼的枕骨、成對的齒骨、許多塊下頜後部的骨頭;頸椎、胸椎、尾椎;肋骨;一個薦骨;成對肩胛骨;成對鳥喙骨;成對肱骨;左橈骨和尺骨;指骨和爪骨;骨盆;左股骨上端和右股骨下端;右腓骨;腳部骨頭包含趾骨。[3][1][7][2]原本的標本編號為BMNH R9951,後來博物館改名重新編號為NHMUK VP R9951。[1][8]

1986年,查理格和米爾納命名了新屬新種沃克氏重爪龍(Baryonyx walkeri),並將BMNH R9951/NHMUK VP R9951標本定為正模標本。屬名由古希臘語的βαρύς/barys(沉重、強壯)和ὄνυξ/onyx(爪子、鉤爪)轉寫組合而來,意即「沉重的爪子」;種名紀念發現正模標本的沃克先生。在當時,敘述者不確定巨爪屬於手部還是腳部,於是根據馳龍科的情況來判斷,暫時假定屬於此類群,但後來才發現是錯的。[9]這隻恐龍曾於同年早些時候在加拿大卓姆赫勒舉辦的一場恐龍系統學研討會上的演講中被介紹過。因為發表的同時化石清修工作仍在持續進行中(當時進度約70%),他們將首篇論文稱作初步性的討論,並承諾之後的日子會再發表更完整的敘述文章。重爪龍是當時全世界所有地方第一種發現的白堊紀早期大型獸腳類恐龍。[1][6]在重爪龍的發現以前,英國最近一次重大的獸腳類發現是1871年的美扭椎龍,因此在一個1986年的訪談中,查理格稱重爪龍是歐洲「本世紀最棒的發現」。[3][4]重爪龍在國際媒體被廣泛報導,記者們以電影《大白鯊》(Jaws)為靈感,將之暱稱為「Claws」。這項發現成為BBC一部1987年紀錄片的主題,並在倫敦自然歷史博物館架設了骨架模型。1997年,查理格和米爾納發表了一份專題研究,更詳細地敘述了正模標本。[3][5][10]重爪龍的正模標本,至今一直是已知最完整的棘龍科骨骼。[11]
額外標本
在英國其他地方以及伊比利半島發現的化石,主要是單獨的牙齒,後來被歸入重爪龍或類似物種。[3]來自懷特島的單獨牙齒和骨頭,包含1998年記載的手部骨頭、2004年英國古生物學家史蒂夫·赫特(Steve Hutt)和潘妮·紐貝瑞(Penny Newbery)所記載的一個脊椎,也被歸入本屬。[12]2017年,英國古生物學家馬汀·蒙特(Martin C. Munt)和同事提到來自懷特島、屬於兩個重爪龍個體的頭骨材料,並表示他們未來會檢驗並敘述這些標本。[13]一個來自西班牙拉里奧哈的上頜骨碎片由西班牙古生物學家路易斯·維耶拉(Luis I. Viera)和何塞·托雷斯(José Angel Torres)於1995年編入重爪龍,[14]但2004年美國古生物學家湯瑪斯·霍茨和同事指出這也很有可能屬於似鱷龍。[15]1999年,來自西班牙布爾戈斯省嬰兒房沉積層(Sala de los Infantes deposit)的一個眶後骨、鱗狀骨、牙齒、脊椎、手指骨及一個趾骨由西班牙古生物學家卡洛里納·維達特(Carolina Fuentes Vidarte)編成一隻未成熟的重爪龍(雖然有些部位是正模標本沒有的)。[16][17]布爾戈斯省附近的恐龍足跡也被認為屬於重爪龍或其他類似的獸腳類。[18]
2011年,來自葡萄牙大山雀口(Boca do Chapim)帕波山組的ML1190標本,是一個破碎的齒骨、牙齒、脊椎、肋骨、骨盆、一個肩胛骨、一個趾骨,由葡萄牙古生物學家奧克塔維奧·馬諦尤斯和同事編入重爪龍,這是這類動物在伊比利半島發現過最完整的遺骸。這副標本的骨骼部位也可見於較完整的正模標本中(除了中部頸椎以外),且尺寸相近。[19]2018年,英國古生物學家托馬斯·亞登(Thomas M. S. Arden)和同事認為葡萄牙標本不屬於重爪龍,因為齒骨前部沒有大幅上翹。[20]伊比利的一些額外棘龍科遺骸可能屬於重爪龍以外的分類單元,包含2018年由葡萄牙古生物學家伊莉莎白·馬拉法亞(Elisabete Malafaia)和同事所敘述、來自莫雷利亞的巴利沃納女獵龍,牠顯然與北非的似鱷龍及亞洲的魚獵龍較近緣。[21][23]
可能異名
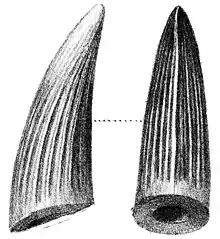
2003年米爾納指出,倫敦自然史博館一些先前被鑑定為鱷龍和斑龍的牙齒可能屬於重爪龍。[7]鱷龍的模式種刀齒鱷龍(Suchosaurus cultridens)由英國生物學家理查·歐文於1841年命名,根據英國地質學家吉迪恩·曼特爾在薩塞克斯郡提爾蓋特森林發現的牙齒。歐文最初認為這些牙齒屬於某種鱷魚;當時他還沒命名及定義出恐龍這個分類群(這是之後幾年的事了)。第二個種吉氏鱷龍(S. girardi)由法國古生物學家亨利·瑟維吉於1897年根據來自葡萄牙大山雀口的下頜碎片和一顆牙齒所命名。2007年法國古生物學家艾瑞克·巴菲陶特認為吉氏鱷龍的牙齒十分類似重爪龍(和刀齒鱷龍),除了高度發育的齒冠稜脊,推測標本屬於同一物種。巴菲陶特認同米爾納認為刀齒鱷龍牙齒與重爪龍幾乎一模一樣的觀點,但表面比較褶皺。前者可能成為後者的首異名,取決於彼此差異是在分類單元內或分類單元間的層級上。根據巴菲陶特所示,有鑑於刀齒鱷龍的正模標本是單一顆牙齒,而沃克氏重爪龍是一副骨骼,保留新的學名是比較切實際的作法。[24][25][26]2011年馬諦尤斯和同事認同鱷龍與重爪龍近緣,但認為前者標本缺乏可鑑定特徵而是個疑名,無法確認其與其他物種的相同性。[19]無論如何,鱷龍都被鑑定為棘龍科,而成為該科第一個命名的物種。[27]
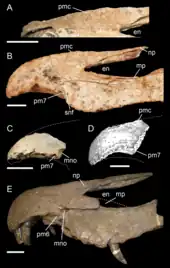
1997年,查理格和米爾納指出,由法國古生物學家菲力普·塔奎特於1984年提及、來自尼日艾爾哈茲組的兩個棘龍科吻部碎片,雖然是來自較年經的阿普第階,但與重爪龍之間有足夠的相似性,因此他們認為這些標本可以被歸類為重爪龍屬下的未定種。[3]1998年,塔奎特和美國古生物學家戴爾·羅素依據這些化石命名為新屬新種拉伯氏脊飾龍(Cristatusaurus lapparenti)。[28]就在同一年稍晚的時候,美國古生物學家保羅·塞里諾和同事根據同樣來自艾爾哈茲組、但較完整的化石,命名發表了另一個新屬新種泰內雷似鱷龍(Suchomimus tenerensis)。2002年,德國古生物學家漢斯-狄特·蘇斯和同事表示,泰內雷似鱷龍與沃克氏重爪龍有足夠的相似性,應該能夠歸類成同一屬下的不同物種:泰內雷重爪龍(Baryonyx tenerensis),而似鱷龍和脊飾龍是同樣的物種。[29]2003年,米爾納認同尼日的化石與重爪龍沒有差異。[7]在一份2004年的研究摘要中,赫特和紐貝瑞支持來自懷特島的大型獸腳類脊椎為重爪龍的異名,並歸類成重爪龍和似鱷龍的近緣物種。[30]後續的研究將重爪龍和似鱷龍維持分開的屬,而有些認為脊飾龍是疑名、或可能與前兩者有些差異。[19][31][32][33][34]2017年,一份由巴西古生物學家卡洛斯·羅伯托·坎德羅(Carlos Roberto A. Candeiro)和同事發表的綜述文章表示,這些物種歸類討論比較偏向語義學而非科學上的辯證,因為普遍認同重爪龍和似鱷龍是各自獨立、但彼此有親緣關係的屬。[35]
敘述

重爪龍估計身長介於7.5至10公尺之間,臀高2.5公尺,體重介於1.2至1.9噸。沃克氏重爪龍的正模標本(NHM R9951)頭骨和脊柱並沒有縫線癒合的跡象,代表這個個體尚未完全成長,因此成熟的個體體型可能更大(如同某些其他棘龍科的情況)。此外,標本的胸骨癒合,表示牠至少在某些程度上已經成熟。[3][36][37][38]
頭骨


發現的重爪龍頭骨不完整,中後部大部分都沒有保存下來。根據與相關物種似鱷龍的比較(約大了20%),估計頭骨全長約91至95公分,。[3][33]頭骨加長,前上頜骨前17公分構成一個長、狹窄、低矮的吻突,上表面光滑圓潤。[3]外鼻孔長且低矮,位於吻尖後方很遠處。吻部前13公分擴張呈湯匙狀,末端形狀呈玫瑰花座狀,類似現代恆河鱷的吻突形狀。前上頜骨下緣前7公分下彎(或鉤狀),而上頜骨前部上翹,這樣的形狀使得下部上排齒列呈S形緣,其中上頜骨前部牙齒朝前突出。吻部在花座狀後方變得特別狹窄;這區域與下頜骨的大顆牙齒相互契合。重爪龍的前上頜骨和上頜骨以複雜的關節互相連接,造成上下頜之間的間隙,稱為吻下凹槽。[3]下彎的前上頜骨、上齒排的S形下緣也可在親緣關係很遠的獸腳類如雙冠龍上觀察到。吻部有許多生前作為血管和神經輸出管道的開孔,[3]而上頜骨似乎有鼻竇的構造。[3][33][37]

重爪龍有個初級次生腭,類似鱷魚,但不類似其他獸腳類。[39]略微皺褶的表面代表嘴巴頂部有個角質墊。鼻骨癒合的特徵將重爪龍與其他棘龍科區別開來,另外眼部上方有道矢狀嵴,位於鼻骨上中線處。嵴狹窄、呈三角形、前部尖銳,末端的十字突朝向後方,這點與其他棘龍科不同。眼睛前的淚骨大致構成一個角,為實心且幾乎呈三角形,類似異特龍,但與其他棘龍科不同。枕骨狹窄,副枕突水平指向外,基翼突加長,向遠延伸到基枕骨(枕骨最下方的骨頭)之下。[3][37][40]塞里諾和同事認為重爪龍的一些頭骨材料被查理格和米爾納鑑定錯誤了,造成枕骨被重建得太深,因此頭骨可能實際上低矮、長而狹窄得像似鱷龍一樣。[40]齒骨前14公分朝著吻部彎曲處上傾。齒骨非常長而淺,內側有明顯的梅克氏凹槽。下頜骨逢(兩半下頜骨前方連接處)特別短。下頜其餘部分纖弱;後部三分之一比前部纖細,外觀呈刀片狀。齒骨前部上彎以容納大科的前齒,此區域並形成下頜的花座狀部分。齒骨和吻部一樣有許多孔洞。[3][33]

大部分與正模標本一起發現的牙齒都沒有與頭骨關節連接;上頜剩少許幾顆,而下頜只剩小的替換牙齒。牙齒呈反曲錐形狀,側面略為變平,曲率幾乎均勻。齒根非常長,朝末端傾斜。[3]齒突(銳利的前後緣)前側和後側均有細密的鋸齒,延伸到整個齒冠。每毫米約有6至8個鋸齒,比其他大型獸腳類如蠻龍、暴龍還多很多。一些牙齒有褶皺,沿內側長面有6至8個稜嵴和細顆粒的琺瑯質(牙齒最外層),而其他牙齒則沒有;這些存在可能與位置或成長發育有關。[3][33]每個齒列內側都有個骨質壁。與大多數其他獸腳類相比牙齒的數量非常多,每個前上頜骨有6至7顆,每個齒骨有32顆。根據緊密排列及較小的齒骨牙齒與前上頜骨對應長度的牙齒相比,上下頜牙齒數量間的差異似乎比其他獸腳類要來得更明顯。[3]正模標本上頜末端的花座狀有13個齒槽(左6右7)顯示兩邊牙齒數量不對稱。前四顆上排牙齒較大(其中第二、三顆最大),第四、五顆逐漸變小。[3]最大直徑為最小的兩倍。齒骨前四個齒槽(對應上頜尖端)最大,其餘尺寸平均。齒槽間有亞三角形的齒內板。
顱後骨骼

雖然重爪龍的頸部起初被認為缺乏獸腳類典型的S型曲線,[3]實際上頸部看起來確實有呈現S形,雖然還是比其他獸腳類要直得多。[41]頸椎向頭部逐漸變細、由前到後逐漸變長。其關節突平坦(連接頸椎的突起),而上骺(頸部肌肉附著的突起)發育良好。樞椎(第二頸椎)與頭骨相比較小,並有發育良好的下椎弓突(hyposphene)。頸椎神經弓並非全部與椎體癒合,那邊的神經棘低矮而纖細。頸肋短,類似鱷魚,可能某種程度上彼此重疊。背部脊椎椎體尺寸相近。如同其他獸腳類,重爪龍的骨骼呈現氣腔化,藉由神經弓和椎體(主要靠近橫突)側凹的開孔。背部神經棘從前到後由短而實變成高而寬。背部神經棘纖細、中等程度地加長,代表沿背部中央可能有道背脊或隆起的構造(雖然與其他棘龍科相較之下,發育程度僅為初級)。重爪龍薦骨與尾部前段的脊椎兩側明顯凹入,這在棘龍科中相當獨特。[3][6][40]

鳥喙骨輪廓看起來呈錐形向後,與肩胛骨以扣槽相連,這在棘龍科中非常獨特。肩胛骨健壯,與身軀相比前肢骨頭短,但寬而結實。肱骨短而粗壯,末端大幅擴張而平坦,上側為三角肌嵴和肌肉附著,下側連接尺骨和橈骨關節。橈骨短、厚實而筆直,比肱骨長度的一半還短,而尺骨略長一些。尺骨有強壯的肘突和擴張的下端。手部有三指;第一指沿弧線長有測量長約31公分的大指爪,指爪生前可能覆有角蛋白鞘而更長。除了尺寸以外,爪子的比例上在獸腳類中相當典型,即左右對稱、略微收縮、圓潤且尖端鋒利。沿爪子長度方向有一道凹槽延伸。其餘的指爪較小。髂骨(骨盆的主要構成)有明顯的上髖臼嵴(supracetabular crest)、纖細而垂直擴張的前突、長且筆直的後突,還有明顯的短肌架(brevis shelf)和面向下的深凹槽。髖臼(連接股骨的凹槽)的前後很長。坐骨(骨盆下方最遠端)上部有個發育良好的閉孔突。恥骨下端邊緣外轉,恥靴無擴張。股骨缺乏腓髁上的凹槽,且腓骨有非常淺的腓窩,這在棘龍科中相當獨特。[3][37][6][40]
分類

在查理格和米爾納最初的敘述中,他們認為重爪龍有著足夠的獨特性,可以建立成一個新的演化支:重爪龍科(Baryonychidae),並覺得牠不太可能屬於其他任何獸腳類類群;他們同時也考慮了重爪龍實際上可能是種槽齒目(一個原始主龍類集合,現在被認為是個並系群)的可能性,因為牠擁有許多顯著的原始特徵;但又指出前上頜和上頜骨間的關節與雙冠龍之間有相似性。此外,他們還提到兩個來自尼日的吻部化石(後來成為脊飾龍的建立依據),曾於1984年由塔奎特編入棘龍科,顯然與重爪龍最為相似,而應該一起編入重爪龍科。[1]1988年美國古生物專家葛瑞格利·保羅同意塔奎特的觀點,於1915年根據來自埃及的碎片化石建立的棘龍屬(化石已毀於二戰)與重爪龍在彎曲吻部上具有相似性,可能是倖存至最後的雙冠龍科。[38]巴菲陶特也於1989年支持這項親緣理論。[42]1990年,查理格和米爾納反駁重爪龍與棘龍科的關聯性,因為沒發現兩者化石間足夠的相似處。[6]但後來他們於1997年認同了重爪龍科和棘龍科的關聯性,但不認同將前者作為後者的異名,鑒於重爪龍的化石較完整,比破碎的棘龍更適合當作科的模式屬,而且也沒發現彼此有足夠顯著的相似性。[3]2004年,霍茨和同事將重爪龍科列為棘龍科的異名。[15]
1990年代的發現為重爪龍的親緣關係提供了更進一步的線索。1996年,一個來自摩洛哥的吻部化石被歸入棘龍,而來自巴西的激龍和崇高龍(彼此可能是異名)也被命名。[43][29]1998年,根據尼日的化石建立了脊飾龍和似鱷龍。在塞里諾等人敘述似鱷龍的論文中,將前者和重爪龍放入棘龍科內新成立的演化支重爪龍亞科(Baryonychinae),而棘龍和激龍則屬於棘龍亞科(Spinosaurinae)。重爪龍亞科的特徵在於:較小的體型、嘴部末端花座後方的齒骨牙齒數量較多、牙齒邊緣有鋸齒狀、以及前部胸椎深突;棘龍亞科的特徵為:筆直且缺乏鋸齒狀的齒冠、前上頜骨第一齒小顆、頜部中的牙距增加、鼻孔可能位於後面離吻尖遠、高聳的神經棘帆。[40][28]他們還將棘龍科與其近緣物種一起編入棘龍超科(Spinosauroidea),但2010年英國古生物學家羅傑·班森(Roger Benson)認為這是較早命名的斑龍超科(Megalosauroidea)的次異名。[44]在一份2007年的研究摘要中,美國古生物學家丹佛·福勒(Denver W. Fowler)表示既然鱷龍才是科內第一個命名的物種,因此棘龍超科、棘龍科、重爪龍亞科分別都應該以鱷龍超科(Suchosauroidea)、鱷龍科(Suchosauridae)、鱷龍亞科(Suchosaurinae)來取代,無論重爪龍這個學名有無保留。[27]2017年,巴西古生物學家馬可斯·塞爾斯(Marcos A. F. Sales)和西瑟·舒茨(Cesar L. Schultz)認為重爪龍亞科的可靠性不足,因為鋸齒邊緣的牙齒可能是棘龍科的祖徵。[34]

以下演化樹顯示重爪龍在棘龍科中的位置,取自2018年湯馬斯·亞登(Thomas M. S. Arden)和同事的研究:[45]
| 棘龍科 Spinosauridae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
演化

白堊紀巴列姆階至森諾曼階(約1億3000萬至9500萬年前)時,棘龍科似乎分布得相當廣泛,而已知最古老的棘龍科化石可追溯至侏羅紀中期。[46]牠們的共同特徵包含長而狹窄、鱷魚狀的頭骨;半圓形牙齒,邊緣的鋸齒狀從細到無;吻部末端的花座狀;增強耐受力的次生腭。相較之下,原始及典型獸腳類為高而狹窄的吻部、刀片狀的牙齒有鋸齒狀突起。[47]棘龍科與鱷魚相似的頭骨形態適應,是趨同演化的結果;後者的早期物種一開始也具有類似典型獸腳類的頭骨,後來才發展成加長的吻部、圓錐形牙齒和次生腭。這樣的適應可能是因為掠食習慣從獵捕陸生動物轉變成魚類所造成的。但與鱷魚不同的是,棘龍科的顱後骨骼似乎沒有發展出水棲型的適應形態。[48][47]塞里諾和同事於1998年推測棘龍科的大指爪和強壯手臂在侏羅紀中期就發展出來,發生於頭骨變長和其他跟魚食性有關的適應特徵之前,因為前述的特徵是與牠們的斑龍科親戚所共有的。他們還表示棘龍亞科和重爪龍亞科的分化大約發生於白堊紀早期的巴列姆階。[40]
關於棘龍科的古生物地理學已經提出了許多種理論。鑒於北非的似鱷龍和歐洲的重爪龍較近緣,而離同樣來自北非的棘龍較遠,因此棘龍科的分布不能以大陸漂移造成的異域物種形成來解釋。塞里諾和同事提出棘龍科最初分布於盤古超大陸,後來因為特提斯洋而分開。當時棘龍亞科可能在南方(非洲、南美洲:岡瓦納大陸)演化,而重爪龍亞科在北方(歐洲:勞亞大陸),似鱷龍則是北部物種向南部擴散的結果。[40]2002年,巴菲陶特和突尼西亞古生物學家穆罕默德·瓦哈(Mohamed Ouaja)提出重爪龍亞科可能是棘龍亞科的祖先,後者顯然在非洲取代了前者的地位。[49]2003年,米爾納提出棘龍科起源於侏羅紀的勞亞大陸,並透過伊比利亞陸橋演化輻射遷徙到岡瓦納。[7]2007年,巴菲陶特指出古地理學研究顯示白堊紀早期時伊比利亞是靠近北非的,成為歐洲和非洲之間的墊腳石,從而支持米爾納的理論,並由伊比利亞出產的重爪龍亞科得到佐證。歐非間的擴散方向尚未明瞭,[24]且後續在亞洲乃至澳洲發現的棘龍科遺骸代表情況可能更複雜。[19]
2016年,西班牙古生物學家亞歷杭卓·色拉諾-馬丁尼茲(Alejandro Serrano-Martínez)和同事發表了已知最古老的棘龍科化石:來自侏羅紀中期尼日的牙齒,加上其他侏羅紀棘龍科牙齒也都來自非洲,於是推測棘龍科起源於岡瓦那大陸,但後續的散布途徑仍不清楚。[46]2017年,坎德羅和同事提出岡瓦納北部的棘龍科被其他掠食者所取代,如阿貝力龍科,因為自森諾曼階之後全世界任何地方都沒有找到確定的棘龍科化石。他們將棘龍科的消失、以及岡瓦納大陸物種組成的轉換,歸咎於環境變遷,可能是海侵作用所造成。[35]2020年,馬拉法亞和同事表示重爪龍依然是最古老的已確認棘龍科,同時認同其他較古老的遺骸也被暫時歸類為棘龍科。[23]
古生物學

食性和覓食習慣
1986年,查理格和米爾納提出,加長的吻部、許多細鋸齒狀牙齒代表重爪龍是種食魚動物,推測牠蹲在河岸邊使用爪子作為魚叉將魚抓出水面(類似現代的灰熊)。[1]兩年後,塔奎特[50]指出,尼日的棘龍科吻部近似現代的恆河鱷,並推測有著近似鷺科和鸛科的習性。[3][1]1987年,蘇格蘭古生物學家安德魯·齊臣納(Andrew Kitchener)反駁重爪龍的食魚習性,並提出是食腐動物的可能性,使用長頸子在地面掠食、爪子扒開屍體、長吻部(鼻孔位於後部以便呼吸)深入獵物體腔來探勘。[51]齊臣納表示重爪龍的頜部和牙齒用來殺死其他恐龍顯得太脆弱,且有著太多食魚性的適應性構造,用來抓魚又顯得太厚重。[51]根據愛爾蘭古生物學家羅賓·雷德(Robin E. H. Reid)所述,一個被腐食的屍體能被其掠食者所分解,而能這麼做的大型動物(如灰熊)也有能力捕魚(至少在淺水中)。[52]

1997年,查理格和米爾納在沃克氏重爪龍正模標本胃部發現了直接的食物證據:被胃酸侵蝕的魚鱗和牙齒,屬於鱗齒魚的曼氏申斯蒂魚,成為獸腳類中第一件魚食性證據。[53]此外,還在胃部發現了年輕禽龍類已磨損的骨頭。另外還有發現魚食性的間接證據,像是用來捕抓和吞下獵物、形似鱷魚的適應性構造:長而狹窄的頜部和末端的花座狀、以及吻部下彎的尖端及凹槽,形似恆河鱷。在他們看來,這些適應構造能使重爪龍用像鱷魚一樣的方式來捕捉中小型魚類:以吻部的凹槽抓住魚類(使牙齒具備刺穿功能),再將頭部仰起,然後吞下。[3]較大隻的魚則可以先用爪子撕碎。下頜牙齒比上頜牙齒更小、更密集且數量更多,可能有助於抓取食物。查理格和米爾納提到重爪龍可能主要以魚類為食(雖然牠可能也是活躍的掠食者或投機的食腐者),但並沒有成為像異特龍那樣頂級掠食者的條件。他們認為重爪龍以前肢和大爪捕捉、殺死及撕開較大型的獵物。標本中還發現了明顯的胃石(砂囊石)。[3]德國古生物學家奧利佛·溫斯(Oliver Wings)於2007年認為在獸腳類(如重爪龍和異特龍)中找到的少量胃石是意外被吞下的。[54]巴菲陶特和同事於2004年的研究中提到,在一個巴西的翼龍類頸椎發現嵌有一顆棘龍科牙齒,確認了後者並非僅是魚食性的。[55]
2007年,英國古生物學家艾蜜莉·瑞菲德(Emily J. Rayfield)和同事使用電腦斷層掃描檢驗吻部進行有限元素分析,指出重爪龍的生物力學與恆河鱷最相似,比較不像美國短吻鱷和其他普通獸腳類,支持了棘龍科魚食性的理論。[39]2013年,英國古生物學家安德魯·庫夫(Andrew R. Cuff)和瑞菲德的棟樑理論研究將CT掃描的棘龍科吻部的生物力學與現代鱷魚作比較,發現重爪龍和棘龍吻部對於承受彎曲和扭轉動作產生的壓力的耐受力相似。重爪龍被發現有著比棘龍和恆河鱷還高的前後向吻部耐受力。研究人員得出結論(與2007年的研究結果相反):重爪龍的檢驗結果與恆河鱷不同;棘龍科並非單一依賴魚食性維生的動物,其食性取決於個體體型。[8]加拿大古生物學家法蘭斯華·泰利恩(François Therrien)和同事先前於2005年的棟樑理論研究未能重建重爪龍的力學數據,但發現近緣的似鱷龍可能使用頜部前部來抓住獵物,並推測棘龍科的頜部特化用來獵捕除了魚類之外還有較小型的陸生獵物。他們猜想棘龍科可以用花座狀結構的前端牙齒固定小獵物,並透過將其甩動來殺死。較大型的獵物則不是由嘴部撕咬,而是用前肢來捕捉和殺死,因為頭骨無法承受大動作的壓力或衝擊。他們亦認同棘龍科的圓錐形牙齒發育良好可以刺穿並固定獵物,形狀使其能夠承受不同方向動作的壓力。[56]
2014年,在美國古生物學家丹尼·安度薩(Danny Anduza)和福勒的研究摘要中,指出棕熊並不像重爪龍推測的那樣抓魚,並排除恐龍會像蒼鷺那樣把頭部快速射向水中的可能性,因為棘龍科的頸部沒有大幅的S曲線,且眼睛位置不佳,無法構成良好的雙眼視覺。於是他們推測重爪龍先將頜部用橫撈的方式來抓魚(如同恆河鱷),搭配指爪來固定及刺穿大隻的魚,最後再用頜部進行控制(類似棕熊和漁貓)。他們不認為棘龍科牙齒適合分解獵物(因為缺乏鋸齒狀),也不能用指爪分解,而可能把整個獵物直接吞下。[57]2016年,比利時古生物學家克里斯托福·亨德里克斯(Christophe Hendrickx)和同事的研究發現成年棘龍科在下頜受到壓力時可以側向移動下頜的兩邊分支,從而使咽部(嘴部通往食道的開口)變寬。這種頜部關節也可在翼龍類及現存的鵜鶘觀察到,並同樣使棘龍科能夠吞下大型獵物,如魚類和其他動物。他們還提到葡萄牙可能的重爪龍化石與單獨的禽龍牙齒一同被發現,將這與其他類似的證據一起列出,來支持棘龍科的投機性掠食行為。[33]同年另一個由法國古生物學家羅曼·弗洛(Romain Vullo)和同事的研究發現棘龍科頜部與海鰻科是趨同演化;這類魚類也同樣擁有側向收縮的頜部(而鱷魚頜部則是縱向收縮)、加長的吻部和帶有加大牙齒的花座狀末端、及花座後方凹槽帶有較小的牙齒。這樣的頜部可能是演化來在光線較暗的水域環境捕捉獵物,且可能有助於偵測獵物。[58]
運動和水生棲地

在查理格和米爾納最初的敘述中,他們沒有考慮重爪龍為水生(因為鼻孔位於吻部兩側遠離前端,加上顱後骨骼的形態),但認為牠具有游泳能力,如同大部分陸棲脊椎動物。[1]他們推測加長的頭骨、長頸部、以及強健的肱骨代表重爪龍為半四足步行,在獸腳類中相當特殊。[1]在他們1997年的論文中,發現沒有骨骼上的證據來支持這點,但維持認為前肢可能強壯到足以維持四足姿勢,並可能在蹲(或四腳著地)於水面附近(或水中)時能夠抓起水生獵物。[3]2014年,德國-摩洛哥古生物學家尼薩·伊卜拉辛和同事根據新標本所作的棘龍重新敘述論文中,基於身體重心前傾,推測牠是四足動物。作者並認為四足行走對於重爪龍來說是不太可能的,因為依照近緣的似鱷龍較完整的腿部來看,並不支持這樣的姿勢。[48]已經提出過許多關於棘龍科高神經棘帆的理論,例如用來作體溫調節、類似駝峰構造的脂肪儲存、或展示功能,而在2015年,德國生物物理學家揚·金薩(Jan Gimsa)和同事提出這項結構可能有利於水中運動,當潛游時增進操控度,並當作頸部與尾部大幅活動時的支點,類似現存的旗魚和長尾鯊。[59][60]
2017年,英國古生物學家大衛·宏恩(David E. Hone)和霍茨假設棘龍科的頭冠可能用於性別或威嚇展示。作者們亦指出沒有理由相信重爪龍的前肢有能力旋轉(經過下臂尺橈骨來旋轉手掌),從而依靠手掌支撐或行走。支撐或使用前肢進行移動是有可能的(如停止狀態的獸腳類足跡所示),但若這是平常在作的事,前肢應該對此會顯示出適應跡象。宏恩和霍茨進一步表示棘龍科的前肢對於抓捕獵物似乎不是最有效率的功能,反而看起來與擅於挖掘的動物前肢很相似。他們推測挖掘能力在開鑿巢穴、挖洞找水、或伸向某些獵物時會很有用處。宏恩和霍茨也相信棘龍科會涉行或浸行水中,而非將整個身體潛入游泳,因為水棲性的適應構造表現得並不明顯。[11]2018年,加拿大古生物學家唐納·韓德森(Donald M. Henderson)進行的浮力研究發現,關係較遠的其他獸腳類漂浮程度與受檢驗的棘龍科一樣,從而支持牠們可能待在水濱或淺水,而非半水棲。[61]
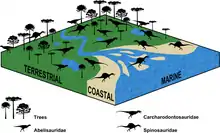
2010年,法國古生物學家羅曼·阿米特(Romain Amiot)和同事將世界各地棘龍科牙齒與其他獸腳類及現代動物間的氧同位素組成進行比較,提出棘龍科為半水生。棘龍科可能花費一天的大部分時光待在水中,就像鱷魚和河馬,並與前者有類似的食性;兩者都是機會主義的掠食者。鑒於大部分棘龍科似乎沒有為水生而適應出的解剖學構造,作者推測潛入水中是種體溫調節的展現,鱷魚、河馬也有相同的行為。棘龍科轉變成水棲和魚食性可能也是為了避免與其他大型陸棲型獸腳類競爭而作出的棲位分化。[62]2016年,塞爾斯和同事對棘龍科、阿貝力龍科、鯊齒龍科的化石分布進行了統計分析,結果發現棘龍科與水岸環境有著最密切的關聯性支持。棘龍科似乎也棲息在內陸環境(分布與鯊齒龍科相近),代表牠們對於棲地的接受度比普遍認為的更廣泛。[63]
2017年塞爾斯和舒茨根據頭骨結構,如圓錐形牙齒、側扁的吻部、位置後退的鼻孔,認同棘龍科為半水生及部分魚食性。他們解釋道,組織學數據表明,棘龍科部分物種比其他的更偏於陸棲性,反映出牠們之間的生態棲位分化。由於棘龍科某些物種的鼻孔比其他物種的小,推測其嗅覺能力較弱,這也可在現代魚食性動物身上觀察到,而牠們捕魚時可能依賴其他種感官(如視覺或力學感受器)的協助。嗅覺可能對也會獵捕陸生動物的棘龍科更有幫助,如重爪龍亞科。[34]2018年,法國古生物學家奧古斯特·哈瑟勒(Auguste Hassler)和同事檢驗北非獸腳類牙齒內含的鈣同位素研究,發現棘龍科有著同時獵捕魚類和植食性恐龍的混合食性,而其他獸腳類(阿貝力龍科和鯊齒龍科)的檢驗結果則主要獵捕植食性恐龍。這證實了這些獸腳類間的棲位分化現象,並且棘龍科是半水生掠食者。[64]
2017年,德國古生物學家卡加·華斯科(Katja Waskow)和馬諦尤斯的生長線組織學研究,發現可能的葡萄牙重爪龍標本死亡時間介於23至25歲,並已接近最大體型和骨骼成熟。這與神經中樞縫線未癒合所暗示的較小年齡互相矛盾,成熟和亞成年特徵同時存在可能是幼態延續(幼年特徵維持到成年)所造成。幼態延續特徵也許與游泳運動有關聯,因為這些特徵也在其他被認為是水生的已滅絕動物(如蛇頸龍類和離片椎目)身上觀察到。研究還發現這隻動物在13至15歲左右時達到性成熟,是根據這段時間的生長線減緩來判定。[65]2018年,巴西古生物學家提托·奧雷里亞諾(Tito Aureliano)和同事記載了一個來自巴西的棘龍科脛骨顯示出骨頭的高密度,這是一項與半水生四足類有關的特徵,用來增加身體密度,以降低因肺部吸氣產生的浮力;其中半水生哺乳類的高密度骨頭是用來適應生活在淺水環境。[66]
古生態學

威爾德黏土組由豪特里維階(下部)至巴列姆階(上部)時期的沉積層構成,約為1億3000萬至1億2500萬年前。沃克氏重爪龍的正模標本發現於後者非海洋的靜止水域泥土中,這裡被復原成一個充滿淺水、潟湖和草沼的河流兼泥灘環境。[3]早白堊世的威爾德地區(薩里郡、薩塞克斯郡、肯特郡),由大片淡水至汽水的威爾登湖所覆蓋。兩條大河流疏通北部地區(相當於現在的倫敦),通過三角洲流進湖泊;盎格魯-巴黎盆地位於其南部。氣候屬亞熱帶,類似現在的地中海盆地。由於煙囪坑由許多層級的地層組成,在這裡發現的物種生前不一定同時生活過。[3][67][68]同地點的恐龍包含鳥腳類的曼特爾龍、禽龍、以及小型蜥腳類。[69]威爾德黏土組的其他脊椎動物有鱷魚、翼龍類、蜥蜴(如多塞特龍)、兩棲類、鯊魚(如弓鮫)、以及硬骨魚(包括申斯蒂魚)。[70][71]無脊椎動物有介形綱、等足目、蚌蝦、雙殼綱、還鑑定出至少十個昆蟲類群的物種(包括Valditermes、Archisphex、Pterinoblattina等)。[72][73]植物中,威西利亞蕨和水生草本植物貝氏草(Bevhalstia)相當普遍。其他植物發現還有蕨類、木賊、石松、毬果植物。[74][75]
懷特島威塞克斯組的其他恐龍包含:獸腳類的新獵龍、極鱷龍、鞘虛骨龍、簧椎龍、聯鳥龍;鳥腳類的禽龍、稜齒龍、荒漠龍;蜥腳類的鳥面龍、優腔龍、軟骨龍;甲龍類的多刺甲龍。[76]葡萄牙發現可能屬於重爪龍標本的帕波山組由泥灰岩構成,代表曾是潟湖環境;這邊發現的其他恐龍遺骸有暫時編入曼特爾龍的化石碎片、一種大鼻龍類的蜥腳類、以及斑龍類。[19][24]
埋藏學

查理格和米爾納提出了一種可能的情境來解釋沃克氏重爪龍正模標本的埋藏過程(屍體被分解及化石化的變化過程)。[3]骨骼周圍的細顆粒沉積物,加上骨骼被發現位置分布彼此接近(頭骨和前肢位在挖掘區域的一端,而骨盆和後肢則位在另一端),代表在沉積作用當時的環境是平靜的,水流並未將屍體沖到遠處,可能因為水很淺的關係。標本死亡地點似乎與身為魚食性動物很相符合。這隻動物可能已經抓到魚並在泥濘平原上享用,之後陷入泥潭然後死亡被掩埋。由於骨骼保存完好、沒有咬痕,屍體似乎不曾被食腐動物所破壞(並推測很短時間就被沉積物掩埋)。[3]
骨骼關節脫落可能是軟組織分解造成的。骨骼某些部分已經歷不同程度的風化作用,可能是因為水位或沉積物發生變化,使得骨骼部分暴露在外所造成。肢帶、肢骨、齒骨及肋骨都在化石化之前就破碎了,可能是掩埋時遭到大型動物踩踏的結果。尾巴的大部分似乎在化石化之前就已遺失,也許是被食腐清掉、或腐爛漂走了。骨頭方向表明屍體在背側躺下(可能稍微左傾、右側朝上),可用來解釋為何所有下頜牙齒都脫離齒槽、而有些上頜牙齒依然留在原位。[3][2]葡萄牙標本ML1190的大部分骨骼受損,一些刮痕可能由小型食腐動物造成。標本關節脫落代表是在較偏陸地的環境被移動(遺失很多骨骼),但剩下的骨骼被發現時位置彼此靠近。[19][24]
參考來源
- Charig, A. J.; Milner, A. C. . Nature. 1986, 324 (6095): 359–361. Bibcode:1986Natur.324..359C. PMID 3785404. doi:10.1038/324359a0.
- Psihoyos, L.; Knoebber, J. . London: Cassell. 1994: 176–179. ISBN 978-0679431244.
- Charig, A. J.; Milner, A. C. . Bulletin of the Natural History Museum of London. 1997, 53: 11–70 [2021-01-14]. (原始内容存档于2014-08-08).
- Edwards, D. D. . Science News. 1986, 130 (23): 356. JSTOR 3970849. doi:10.2307/3970849.
- Moody, R. T. J.; Naish, D. . Geological Society, London, Special Publications. 2010, 343 (1): 89–109. Bibcode:2010GSLSP.343...89M. doi:10.1144/SP343.6.
- Charig, A. J.; Milner, A. C. . Carpenter, K.; Currie, P. J. (编). . Cambridge: Cambridge University Press. 1990: 127–140 [2021-01-14]. ISBN 978-0-521-43810-0. doi:10.1017/CBO9780511608377.012. (原始内容存档于2021-04-14).
- Milner, A. C. . Actas de las II Jornadas Internacionales Sobre Paleontologýa de Dinosaurios y Su Entorno. 2003: 129–138.
- Cuff, A. R.; Rayfield, E. J. Farke, Andrew A , 编. . PLOS ONE. 2013, 8 (5): e65295. Bibcode:2013PLoSO...865295C. PMC 3665537
 . PMID 23724135. doi:10.1371/journal.pone.0065295.
. PMID 23724135. doi:10.1371/journal.pone.0065295. - Norman, D. B. . . New York: Crescent Books. 1985: 57–58. ISBN 978-0-517-46890-6.
- Osterloff, E. . www.nhm.ac.uk. 2018 [20 October 2018]. (原始内容存档于2021-04-14).
- Hone, D. W. E.; Holtz, T. R. . Acta Geologica Sinica – English Edition. 2017, 91 (3): 1120–1132 [2021-01-14]. doi:10.1111/1755-6724.13328. (原始内容存档于2019-07-19).
- Clabby, S. M. . DinoWight. 2005 [12 October 2015]. (原始内容存档于2018-10-10).
- Munt, M. C.; Blackwell, G.; Clark, J.; Foster, B. . SVPCA. 2017, 65: 1. doi:10.13140/RG.2.2.17925.86242.
- Viera, L. I.; Torres, J. A. . Munibe Ciencias Naturales. 1995, 47: 57–61 [2021-01-14]. ISSN 0214-7688. (原始内容存档于2021-04-14) (西班牙语).
- Holtz, T. R.; Molnar, R. E.; Currie, P. J. . Weishampel, D. B.; Dodson, P.; Osmolska, H. (编). 2. Berkeley: University of California Press. 2004: 71–110. ISBN 978-0-520-24209-8.
- Vidarte, C. F.; Calvo, M. M.; Meijide, M.; Izquierdo, L. A.; Montero, D.; Pérez, G.; Torcida, F.; Urién, V.; Fuentes, F. M.; Fuentes, M. M. . Actas de las I Jornadas Internacionales Sobre Paleontología de Dinosaurios y Su Entorno. Salas de los Infantes, Burgos. 2001: 349–359 (西班牙语).
- Pereda-Suberbiola, X.; Ruiz-Omeñaca, J. I.; Canudo, J. I.; Torcida, F.; Sanz, J. L. . Godefroit, P. (编). . Indiana University Press. 2012: 389–390 [2021-01-14]. ISBN 978-0-253-00570-0. (原始内容存档于2021-04-14).
- Pérez-Lorente, F. . Life of the Past. Indiana: Indiana University Press. 2015: 325. ISBN 978-0-253-01515-0.
- Mateus, O.; Araújo, R.; Natário, C.; Castanhinha, R. (PDF). Zootaxa. 2827. 2011, 2827: 54–68 [2021-01-14]. doi:10.11646/zootaxa.2827.1.3. (原始内容存档 (PDF)于2016-09-19).
- Arden, T. M.S.; Klein, C. G.; Zouhri, S.; Longrich, N. R. . Cretaceous Research. 2018, 93: 275–284. doi:10.1016/j.cretres.2018.06.013.
- Malafaia, E.; Gasulla, J. M.; Escaso, F.; Narváez, I.; Sanz, J. L.; Ortega, F. . Cretaceous Research. 2018, 92: 174–183. doi:10.1016/j.cretres.2018.08.006.
- Malafaia, E.; Gasulla, J. M.; Escaso, F.; Narvaéz, I.; Ortega, F. . Journal of Iberian Geology. 2020. doi:10.1007/s41513-020-00138-9.
- Buffetaut, E. . Geological Magazine. 2007, 144 (6): 1021–1025. Bibcode:2007GeoM..144.1021B. doi:10.1017/S0016756807003883.
- Hendrickx, C.; Mateus, O.; Araújo, R. . Journal of Vertebrate Paleontology. 2015, 35 (5): e982797. doi:10.1080/02724634.2015.982797.
- Hendrickx, C. . spinosauridae.fr.gd. 2008 [22 October 2018]. (原始内容存档于2009-12-15) (法语).
- Fowler, D. W. . Journal of Vertebrate Paleontology. 2007, 27 (3): 3 [2021-01-14]. (原始内容存档于2020-03-05).
- Taquet, P.; Russell, D. A. . Comptes Rendus de l'Académie des Sciences, Série IIA. 1998, 327 (5): 347–353. Bibcode:1998CRASE.327..347T. doi:10.1016/S1251-8050(98)80054-2.
- Sues, H. D.; Frey, E.; Martill, M.; Scott, D. M. . Journal of Vertebrate Paleontology. 2002, 22 (3): 535–547. doi:10.1671/0272-4634(2002)022[0535:icasdt]2.0.co;2.
- Hutt, S.; Newbery, P. . Symposium of Vertebrate Palaeontology and Comparative Anatomy. 2004. (原始内容存档于5 October 2015).
- Allain, R.; Xaisanavong, T.; Richir, P.; Khentavong, B. . Naturwissenschaften. 2012, 99 (5): 369–377. Bibcode:2012NW.....99..369A. PMID 22528021. doi:10.1007/s00114-012-0911-7.
- Benson, R. B. J.; Carrano, M. T.; Brusatte, S. L. (PDF). Naturwissenschaften (Submitted manuscript). 2009, 97 (1): 71–78 [2021-01-14]. Bibcode:2010NW.....97...71B. PMID 19826771. doi:10.1007/s00114-009-0614-x. (原始内容存档 (PDF)于2018-11-13).
- Hendrickx, C.; Mateus, O.; Buffetaut, E.; Evans, A. R. . PLOS ONE. 2016, 11 (1): e0144695. Bibcode:2016PLoSO..1144695H. PMC 4703214 . PMID 26734729. doi:10.1371/journal.pone.0144695.
- Sales, M. A. F.; Schultz, C. L. . PLOS ONE. 2017, 12 (11): e0187070. Bibcode:2017PLoSO..1287070S. PMC 5673194 . PMID 29107966. doi:10.1371/journal.pone.0187070.
- Candeiro, C. R. A.; Brusatte, S. L.; Souza, A. L. . Anuário do Instituto de Geociências – UFRJ. 2017, 40 (3): 294–302. doi:10.11137/2017_3_294_302.
- Therrien, F.; Henderson, D. M. . Journal of Vertebrate Paleontology. 2007, 27 (1): 108–115. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- Paul, G. S. . Princeton University Press. 2010: 87–88. ISBN 978-0-691-13720-9.
- Paul, G. S. . New York: Simon & Schuster. 1988: 271–274. ISBN 978-0-671-61946-6.
- Rayfield, E. J.; Milner, A. C.; Xuan, V. B.; Young, P. G. . Journal of Vertebrate Paleontology. 2007, 27 (4): 892–901. doi:10.1671/0272-4634(2007)27[892:FMOSCD]2.0.CO;2.
- Sereno, P. C.; Beck, A. L.; Dutheil, D. B.; Gado, B.; Larsson, H. C. E.; Lyon, G. H.; Marcot, J. D.; Rauhut, O. W. M.; Sadleir, R. W.; Sidor, C. A.; Varricchio, D. D.; Wilson, G. P.; Wilson, J. A. . Science. 1998, 282 (5392): 1298–1302. Bibcode:1998Sci...282.1298S. PMID 9812890. doi:10.1126/science.282.5392.1298.
- Evers, S. W.; Rauhut, O. W. M.; Milner, A. C.; McFeeters, B.; Allain, R. . PeerJ. 2015, 3: e1323. PMC 4614847 . PMID 26500829. doi:10.7717/peerj.1323.
- Buffetaut, E. . Neues Jahrbuch für Geologie und Paläontologie – Monatshefte. 1989, 1989 (2): 79–87. doi:10.1127/njgpm/1989/1989/79.
- Russell, D. A. . Bulletin du Muséum National d'Histoire Naturelle, Paris, 4e Série, Section C. 1996, 18 (2–3): 349–402.
- Benson, R. B. J. . Zoological Journal of the Linnean Society. 2010, 158 (4): 882–935. doi:10.1111/j.1096-3642.2009.00569.x.
- Arden, T. M. S.; Klein, C. G.; Zouhri, S.; Longrich, N. R. . Cretaceous Research. 2018, 93: 275–284. doi:10.1016/j.cretres.2018.06.013.
- Serrano-Martínez, A.; Vidal, D.; Sciscio, L.; Ortega, F.; Knoll, F. . Acta Palaeontologica Polonica. 2015. doi:10.4202/app.00101.2014.
- Holtz Jr., T. R. . Science. 1998, 282 (5392): 1276–1277. doi:10.1126/science.282.5392.1276.
- Ibrahim, N.; Sereno, P. C.; Dal Sasso, C.; Maganuco, S.; Fabri, M.; Martill, D. M.; Zouhri, S.; Myhrvold, N.; Lurino, D. A. . Science. 2014, 345 (6204): 1613–1616 [2021-01-19]. Bibcode:2014Sci...345.1613I. PMID 25213375. doi:10.1126/science.1258750. (原始内容存档于2020-04-07). Supplementary Information (页面存档备份,存于)
- Buffetaut, E.; Ouaja, M. . Bulletin de la Société Géologique de France. 2002, 173 (5): 415–421. doi:10.2113/173.5.415.
- Taquet, P. . Comptes Rendus de l'Académie des Sciences. 1984, 299: 217–222 (法语).
- Kitchener, A. . Nature. 1987, 325 (6100): 114. Bibcode:1987Natur.325..114K. doi:10.1038/325114a0.
- Reid, R. E. H. . Nature. 1987, 325 (6104): 487. Bibcode:1987Natur.325..487R. doi:10.1038/325487b0.
- López-Arbarello, A. . PLOS ONE. 2012, 7 (7): e39370. Bibcode:2012PLoSO...739370L. PMC 3394768 . PMID 22808031. doi:10.1371/journal.pone.0039370.
- Wings, O. . Acta Palaeontologica Polonica. 2007, 52 (1): 1–16 [2021-01-24]. (原始内容存档于2021-04-14).
- Buffetaut, E.; Martill, D.; Escuillié, F. . Nature. 2004, 429 (6995): 33. Bibcode:2004Natur.429...33B. PMID 15229562. doi:10.1038/430033a.
- Therrien, F.; Henderson, D.; Ruff, C. . Carpenter, K. (编). . Indiana University Press. 2005: 179–230. ISBN 978-0-253-34539-4.
- Anduza, D.; Fowler, D. W. . Geological Society of America Abstracts with Programs. 2014, 46 (5): 12 [2021-01-27]. (原始内容存档于2018-12-15).
- Vullo, R.; Allain, R.; Cavin, L. . Acta Palaeontologica Polonica. 2016, 61 [2021-01-27]. doi:10.4202/app.00284.2016. (原始内容存档于2019-07-23).
- Gimsa, J.; Sleigh, R.; Gimsa, U. . Geological Magazine. 2015, 153 (3): 544–547. doi:10.1017/S0016756815000801.
- Bailey, J. B. . Journal of Paleontology. 2015, 71 (6): 1124–1146. doi:10.1017/S0022336000036076.
- Henderson, D. M. . PeerJ. 2018, 6: e5409. PMC 6098948 . PMID 30128195. doi:10.7717/peerj.5409.
- Amiot, R.; Buffetaut, E.; Lecuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, M. A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; Zhou, Z. . Geology. 2010, 38 (2): 139–142. Bibcode:2010Geo....38..139A. doi:10.1130/G30402.1.
- Sales, M. A. F.; Lacerda, M. B.; Horn, B. L. D.; de Oliveira, I. A. P.; Schultz, C. L.; Bibi, F. . PLOS ONE. 2016, 11 (2): e0147031. Bibcode:2016PLoSO..1147031S. PMC 4734717 . PMID 26829315. doi:10.1371/journal.pone.0147031.
- Hassler, A.; Martin, J. E.; Amiot, R.; Tacail, T.; Godet, F. Arnaud; Allain, R.; Balter, V. . Proceedings of the Royal Society B: Biological Sciences. 2018, 285 (1876): 20180197. PMC 5904318 . PMID 29643213. doi:10.1098/rspb.2018.0197.
- Waskow, K.; Mateus, O. . Comptes Rendus Palevol. 2017, 16 (4): 425. doi:10.1016/j.crpv.2017.01.003.
- Aureliano, T.; Ghilardi, A. M.; Buck, P. V.; Fabbri, M.; Samathi, A.; Delcourt, R.; Fernandes, M. A.; Sander, M. . Cretaceous Research. 2018, 90: 283–295 [2021-01-28]. doi:10.1016/j.cretres.2018.04.024. (原始内容存档于2020-03-05).
- Radley, J. D.; Allen, P. . Proceedings of the Geologists' Association. 2012, 123 (2): 382–385. doi:10.1016/j.pgeola.2011.12.005.
- Radley, J. D. . Geology Today. 2006, 22 (3): 109–118. doi:10.1111/j.1365-2451.2006.00563.x.
- Blows, W. T. . New Mexico Museum of Natural History Bulletins. 1998, 14: 29–38 [2021-01-28]. (原始内容存档于2018-11-19).
- Cook, E.; Ross, A. J. . Proceedings of the Geologists' Association. 1996, 107 (3): 231–239. doi:10.1016/S0016-7878(96)80031-9.
- Milner, A. C.; Evans, S. E. . New Mexico Museum of Natural History Bulletins. 1998, 14: 173–176 [2021-01-28]. (原始内容存档于2018-11-19).
- Nye, E.; Feist-Burkhardt, S.; Horne, D. J.; Ross, A. J.; Whittaker, J. E. . Cretaceous Research. 2008, 29 (3): 417–444. doi:10.1016/j.cretres.2008.01.004.
- Cook, E. . Proceedings of the Geologists' Association. 1997, 108 (1): 49–56. doi:10.1016/S0016-7878(97)80005-3.
- Ross, A. J.; Cook, E. . Cretaceous Research. 1995, 16 (6): 705–716. doi:10.1006/cres.1995.1044.
- Batten, D. J. . Cretaceous Research. 1998, 19 (3–4): 279–315. doi:10.1006/cres.1998.0116.
- Martill, D. M.; Hutt, S. . Proceedings of the Geologists' Association. 1996, 107 (2): 81–84. doi:10.1016/S0016-7878(96)80001-0.