赤狐
赤狐(學名:),又名火狐,是狐的一種。赤狐是最大和分佈最廣泛的狐屬食肉目成員之一,它的足迹遍布北半球,包括北美、歐洲和亞洲的大部分地區,以及北非部分地區。赤狐被國際自然保護聯盟列為無危物种。[1] 隨著人類的擴張,赤狐的分布範圍也隨之擴大,它曾被引入澳大利亞,但是在那裡它被認為對当地的哺乳動物和鳥類有害。鉴于赤狐对澳大利亞原生物种造成的干扰,它被列入了“世界百大外來入侵種”名單。[3]
| 赤狐 化石时期: 中更新世–現今 | |
|---|---|
.jpg.webp) | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 哺乳纲 Mammalia |
| 目: | 食肉目 Carnivora |
| 科: | 犬科 Canidae |
| 属: | 狐屬 Vulpes |
| 种: | 赤狐 V. vulpes |
| 二名法 | |
| Vulpes vulpes | |
| 亞種 | |
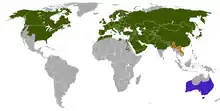 | |
| 赤狐的分佈 原生 引進 不確定是否分布 | |
| 異名 | |
|
Canis vulpes (卡尔·林奈, 1758) | |
赤狐的祖先体型较小,生活于維拉夫蘭期的歐亞大陸[4],並在威斯康辛州冰川融化後不久扩散至北美大陆。[5] 相比狐族其它成员,赤狐在肉食性方面有更多特化[6],除了较大的体型外,它还能更快的适应新环境。虽然名为赤狐,但它也可能发展出其它颜色,包括白化和黑化。[6] 目前已確認有45個赤狐亞種,它们被分為兩類:体型较大的北方赤狐,以及体型较小的,生活于亚洲与北非沙漠地带的南方赤狐基群。[6]
赤狐通常成对或以家庭为单位组成小团体,例如一對伴侶和它們的孩子,或是一只雄赤狐与几只有亲缘關係的雌赤狐。年轻的赤狐伴侣仍需要父母协助照料自己的后代。[7] 赤狐主要以啮齿类为食,但有时也捕食兔子、松鼠、禽类、爬行动物、无脊椎动物和有蹄类幼崽。[8] 它们偶尔也吃水果和植物。[9] 尽管赤狐有能力对付小型捕食者,但它们容易遭受大型捕食者的攻击,如野狼、郊狼、胡狼、大中型猫科动物,以及大型猛禽,例如金雕、雕鸮等都能对它们造成威胁。[10][11]
人类与赤狐的关系由来已久,民间文化与传说中时常有赤狐的形象出现。此外,历史上赤狐被视为有害生物,人们广泛将其猎杀以获取毛皮。因为赤狐分布广泛、数量众多,它一直是皮草贸易的重要目标。[12] 赤狐体型不足以对人类造成威胁,因此随着人类居住地的扩张,赤狐的活动范围也随之变大,并成功适应了许多郊区与城区环境。俄罗斯正在开展赤狐的驯化工作,并成功得到了驯化后的赤狐。
英文名称
.jpg.webp)
在英语中,人们通常把雄性赤狐称為 tods 或 dogs,雌性為 vixens,赤狐幼崽則被稱為 kits。[13] 尽管斯堪的納維亞半島北部分布有少量原生沙狐,而北極狐的分布范围也延伸到了俄羅斯的歐洲部分,但赤狐是唯一原產於西歐的狐狸,因此在口語化的英式英語中,赤狐被簡稱為"the fox"。
演化与分类
_(1980)_Fig._112.png.webp)
赤狐被認為比阿富汗狐、沙狐和孟加拉狐更適應肉食生活。它们的头骨表现出更少的幼态延续,面部肌肉也更加發達。[6] 但赤狐也并非像藏狐那樣以純肉食为生。[6]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
起源
赤狐起源於歐亞大陸,可能從生活于維拉夫蘭中期的 Vulpes alopecoides 或 V. chikushanensis 演化而來。[15] 迄今发现的最早的赤狐化石出土于匈牙利巴兰尼亚州,距今大约340-180万年。[16] 赤狐祖先的体型要比现代物种小。[4] 现代赤狐的化石记录可追溯到更新世中期,与早期人类定居点的垃圾相关。因此有人提出,原始人类可能会猎杀赤狐,将其作为食物或毛皮来源。[17]
扩散
赤狐曾两度扩散至北美大陆,分别在伊利諾伊冰川形成前或发展时,以及威斯康辛州冰川发展时期。[18] 基因图谱显示,北美洲的赤狐与旧世界的同伴在40万年前分离,这或许促进了新物种形成,并与先前的物种 Vulpes fulva 相区分。[19] 在北美洲北部,人们在加拿大梅迪辛哈特的桑加蒙间冰期遗迹中找到了赤狐化石。定年于威斯康辛州冰川时期的化石分别发现于美国多个州的25个不同遗迹,这些州包括:阿肯色、加利福尼亚、科罗拉多、爱达荷、密苏里、新墨西哥、田纳西、德克萨斯、弗吉尼亚和怀俄明。在威斯康辛州冰川时期,赤狐的活动范围延伸到北美南部,而后气候的回暖迫使它们的栖息地向北收缩,直到最近因为人类活动对气候造成的影响,赤狐重新扩散到了南部的土地。[5] 基因检测显示,北美洲存在两个分离于威斯康辛州冰川时期的冰期生物种遗区。北方种遗区位于阿拉斯加与加拿大西部,这里生活有大型赤狐亚种:北阿拉斯加亚种(V. v. alascensis)、英属哥伦比亚省亚种(V. v. abietorum)、北部平原亚种(V. v. regalis)和新斯科舍省亚种(V. v. rubricosa)。南方种遗区(或称为山地种遗区)位于洛矶山脉、喀斯喀特山脉和内华达山脉的亚高山带,这里生活有小型赤狐亚种:喀斯喀特山亚种(V. v. cascadensis)、山地亚种(V. v. macroura)、内华达亚种(V. v. necator)和萨克拉门托河亚种(V. v. patwin)。后一演化支自从末次冰盛期开始独立发展,可能演化出了一些独特的生态与生理适应能力。[18]
尽管欧洲赤狐(V. v. crucigera)自20世纪初期被引入美国部分地区,但最近的基因研究发现,在北美赤狐族群中没有发现任何来自欧洲赤狐的单倍型。[20] 此外,引入美国东部的赤狐已散布至加州的圣华金谷和旧金山湾区,但与萨克拉门托谷的原生赤狐(V. v. patwin)仅在狭窄地带有交集。[21] 目前没有证据表明生活于加州的赤狐与内华达亚种(V. v. necator)或其它山地赤狐(生活于落基山脉和西边的喀斯喀特山脉和内华达山脉之间)有杂交。[22]
亞種
第三版《世界哺乳動物物種》[23]將45個赤狐亞種列為有效物種。2010年,透过單倍型粒線體DNA研究,科学家确定了一个新的赤狐亚种——萨克拉门托河亚种(V. v. patwin),它生活在萨克拉门托谷的草原上。Castello (2018) 区分了旧世界赤狐中的30个亚种以及北美赤狐中的9个亚种。[24]
不同赤狐亚种之间常有基因交流,例如英国赤狐就曾与来自德国、法国、比利时、撒丁岛(或许还包括西伯利亚和斯堪的纳维亚)的赤狐杂交。[25] 不过,基因研究还显示,欧洲各处的赤狐标本中的基因差异非常小。[26][27] 这或许是因为赤狐的活动习性所致,赤狐在一年中的活动范围可达320千米。[28]
歐亞大陸和北非的赤狐亞種分為兩類[6]:
- 北方赤狐,體型大,毛色偏亮。
- 南方沙漠灰狐,包括阿富汗亚种(V. v. griffithi)、白足亚种(V. v. pusilla)和波斯亚种(V. v. flavescens)。这些赤狐的特征介乎北方赤狐与其它狐狸品种之间,它们的头骨更加原始,有更多的幼态延续特征[6],体型也更小。南方沙漠灰狐的最大体型要小于北方赤狐的平均体型,但它们的四肢比例更长,耳朵更大。[6]
| 亚种 | 命名者 | 特征 | 分布范围 | 其它学名 |
|---|---|---|---|---|
| 斯堪的纳维亚亚种 V. v. vulpes
(指名亚种)
|
卡尔·林奈, 1758 | 体型较大,身长70-90cm,体重5-10kg;已知雄性头骨最长为163.2mm。毛发呈现出亮红色,下背部有明显的白色与黄色波纹。[6] | 斯堪的纳维亚半岛,以及前苏联欧洲部分的北部与中部(森林地区),向南延伸到森林草原,东至乌拉尔山脉,向西到欧洲中西部。 | alopex (Linnaeus, 1758) communis (Burnett, 1829) |
| 不列颠哥伦比亚省亚种[29] V. v. abietorum
|
克林顿·哈特·梅里厄姆, 1900 | 总体特征与阿拉斯加北部亚种(V. v. alascensis)相似,但毛发更长,颜色更浅,头骨更加细长。 [30] | 加拿大西北地区的育空地区,以及阿尔伯塔省北部,不列颠哥伦比亚省以及与之交界的美国阿拉斯加州东南海岸。[31] | sitkaensis (Brass, 1911) |
| 阿拉斯加北部亚种V. v. alascensis
|
克林顿·哈特·梅里厄姆, 1900 | 尾巴大且长,耳朵较小,毛发颜色金黄。 [30] | 美国阿拉斯加州安德烈夫斯基荒野 | |
| 外高加索东部亚种 V. v. alpherakyi
|
康斯坦丁·萨杜宁, 1906 | 体型较小,体重大约4kg;雄性头骨最大为132-139mm,雌性则为121-126mm。毛皮呈现出类似铁锈的红色或棕色,沿着脊柱有一条颜色较浅的条纹。被毛短、粗、稀疏。[6] | 土库曼斯坦的盖奥克泰佩,哈萨克斯坦的阿拉尔 | |
| 安纳托利亚亚种 V. v. anatolica
|
奥德菲尔德·托马斯, 1920 | 土耳其的伊兹密尔和爱琴海地区 | ||
| 阿拉伯亚种 V. v. arabica
|
奥德菲尔德·托马斯, 1920 | 阿曼佐法尔省的哈杰尔山脉 | ||
| 阿特拉斯亚种 V. v. atlantica | 约翰·安德烈亚斯·瓦格纳, 1841 | 阿尔及利亚米拉省的阿特拉斯山脈 | algeriensis (Loche维克多·洛奇, 1858) | |
| 拉布拉多亚种 V. v. bangsi
|
克林顿·哈特·梅里厄姆, 1900 | 与北美亚种(V. v. fulva)相似,但耳朵更小,耳朵和腿部的斑纹不明显。[30] | 加拿大拉布拉多地区贝尔岛海峡附近 | |
| 巴巴里亚种 V. v. barbara | 乔治·肖, 1800 | 非洲西北部巴巴里海岸 | acaab (Cabrera, 1916) | |
| 堪察加亚种 V. v. beringiana
|
亚历山大·冯·米登多夫, 1875 | 体型较大,该亚种的毛发是旧世界赤狐中最鲜艳的,呈现出亮红色,背部和侧面的浅色波纹几乎不可见。背毛蓬松柔软。[6] | 西伯利亚东北部的白令海峡附近 | |
| 喀斯喀特山亚种 V. v. cascadensis
|
克林顿·哈特·梅里厄姆, 1900 | 短尾小齿亚种,毛发为黄色而非褐色;它是最有可能产生颜色“交错”的亚种。 [30] | 美国华盛顿州斯卡梅尼亚县的喀斯喀特山脉 | |
| 高加索亚种 V. v. caucasica
|
Dinnik, 1914 | 体型较大,被毛颜色多变,从淡红色到红灰色,再到近乎灰色,毛发短而粗糙。这个亚种可能是由草原亚种(V. v. stepensis)和蒙新亚种(V. v. karagan)的杂交形成的。.[6] | 俄罗斯弗拉季高加索附近 | |
| 欧洲亚种 V. v. crucigera
|
约翰·马托伊斯·贝希斯坦, 1789 | 体型中等,毛发呈黄褐色或红褐色,背部没有白色条纹。[32] 它与斯堪的纳维亚亚种(V. v. vulpes)的主要区别在于它的体型略小,牙齿更小,前臼齿间距较宽。英国的赤狐(也包括澳大利亚的赤狐)常被归为此亚种,尽管其中的许多族群显示出远超欧洲大陆同伴的牙齿密实度。[7] | 欧洲除斯堪的纳维亚半岛、伊比利亚半岛和一些地中海岛屿外的所有赤狐。它也被引入了澳大利亚和北美。 | alba (Borkhausen, 1797) cinera (Bechstein, 1801) |
| 东北亚种 V. v. daurica | 谢尔盖·奥格涅夫, 1931 | 体型较大,沿脊柱的毛发呈鲜艳的黄红色,伴随有白色波纹状条纹,四肢前侧有灰色纵向条纹。被毛粗且蓬松。 [6] | 西伯利亚恰克图以北约45km | ussuriensis (Dybowski, 1922) |
| 纽芬兰亚种 V. v. deletrix | 欧南·班, 1898 | 毛皮总体颜色很浅,在某些部位颜色略深,呈现出金黄或水蓝色。尾巴上没有常见黑色的黑色基地斑点,后脚与爪子很大。[30] | 加拿大纽芬兰岛圣乔治湾 | |
| 俄罗斯远东亚种 V. v. dolichocrania | 谢尔盖·奥格涅夫, 1926 | 西伯利亚乌苏里江南部的Sidemi | ognevi (Yudin, 1986) | |
| 内华达州亚种 V. v. dorsalis | 约翰·爱德华·格雷, 1838 | |||
| 波斯亚种 V. v. flavescens
|
约翰·爱德华·格雷, 1838 | 体型较小,头部类似幼崽,全身覆盖有灰色皮毛。体长49-57.5cm,体重2.2-3.2kg。[6] | 伊朗北部 | cinerascens (Birula, 1913) splendens (Thomas, 1902) |
| 北美亚种 V. v. fulva
|
安塞尔姆·加埃唐·德马雷, 1820 | 体型比斯堪的纳维亚亚种(V. v. vulpes)更小,面部更加细长,尾巴短,毛皮颜色浅并混有白色,腿部毛皮呈深色。[30] | 加拿大和美国东部 | pennsylvanicus (Rhoads, 1894) |
| 阿富汗亚种 V. v. griffithi
|
爱德华·布莱思, 1854 | 比西藏亚种(V. v. montana)略小,毛皮呈现出灰白或银白色。[33]:121 | 阿富汗坎大哈 | flavescens (Hutton, 1845) |
| 科迪亚克亚种 V. v. harrimani
|
克林顿·哈特·梅里厄姆, 1900 | 体型较大,有粗大的尾巴,尾部与下背部皮毛与狼类似。脖子和肩膀上的毛发被拉长并形成了一个褶皱。[30] | 美国阿拉斯加科迪亚克岛 | |
| 华南亚种 V. v. hoole | 郇和, 1870 | [34] | 中国福建省厦门附近 | aurantioluteus (Matschie, 1907) lineiventer (R. Swinhoe, 1871) |
| 撒丁岛亚种 V. v. ichnusae | 格里特·史密斯·米勒, 1907 | 体型小,耳朵也较小。[32] | 意大利撒丁岛,可能也被引入了英格兰中部地区。[25]:6 | |
| 塞浦路斯亚种 V. v. indutus | 格里特·史密斯·米勒, 1907 | 塞浦路斯 | ||
| 雅库茨克亚种 V. v. jakutensis
|
谢尔盖·奥格涅夫, 1923 | 体型较大,但小于堪察加亚种(V. v. beringiana),背部、颈部和肩部毛皮呈现出棕锈色,侧边是更浅的赭黄色。 [6] | 西伯利亚东部雅库茨克以南的针叶林 | sibiricus (Dybowski, 1922) |
| 日本亚种 V. v. japonica
|
谢尔盖·奥格涅夫, 1923 | 日本(不包含北海道) | ||
| 蒙新亚种 V. v. karagan
|
約翰·埃克斯勒本, 1777 | 体型小于斯堪的纳维亚亚种(V. v. vulpes),毛发短而粗,呈现出沙黄色或黄灰色。[6] | 吉尔吉斯斯坦草原 | ferganensis (Ognev, 1926) melanotus (Pallas, 1811) |
| 基奈亚种 V. v. kenaiensis | 克林顿·哈特·梅里厄姆, 1900 | 北美洲体型最大的赤狐亚种之一,毛发比科迪亚克亚种(V. v. harrimani)更软。 [30] | 美国阿拉斯加基奈半岛 | |
| 库尔德斯坦亚种 V. v. kurdistanica
|
康斯坦丁·萨杜宁, 1906 | 体型介于外高加索东部亚种(V. v. alpheryaki)和高加索亚种(V. v. caucasica)之间,皮毛呈浅黄或浅灰色,有时也呈棕红色,比另外两个高加索亚种的毛发更蓬松、密集。 [6] | 土耳其东北部 | alticola (Ognev, 1926) |
| 山地亚种 V. v. macroura
|
斯宾塞·富勒顿·贝尔德, 1852 | 该亚种与北美亚种(V. v. fulvus)类似,但尾巴要长许多,后脚更大,四肢颜色更暗。[30] | 美国犹他州大盐湖附近的瓦萨奇山脉,以及落基山脉的科罗拉多州、犹他州、怀俄明州、蒙大拿州部分,以及加拿大阿尔伯塔省南部 | |
| 西藏亚种 V. v. montana
|
奥利弗·佩恩·皮尔森, 1836 | 相比斯堪的纳维亚亚种(V. v. vulpes),该亚种体型更小,头骨与牙齿也对应比例的更小,毛发较粗。脚底混合有柔软的毛发。[33]:111 | 喜马拉雅山脉 | alopex (Blanford, 1888) himalaicus (Ogilby, 1837) |
| 内华达亚种 V. v. necator
|
克林顿·哈特·梅里厄姆, 1900 | 外表类似北美亚种(V. v. fulvus),尾巴更短,头骨与山地亚种(V. v. macroura)类似。 [30] | 位于加利福尼亚州的内华达山脉高山地带 | |
| 埃及亚种 V. v. niloticus
|
艾蒂安·若弗鲁瓦·圣伊莱尔, 1803 | 体型较小,体长在76.7-105.3cm,尾长30.2-40.1cm,体重1.8-3.8kg。体上方呈红褐色至灰褐色,颈后颜色较深。侧边毛发灰色带有浅黄。[35] 体型比阿拉伯亚种(V. v. arabica)和巴勒斯坦亚种(V. v. palaestina)更大。 | 埃及 | aegyptiacus (Sonnini, 1816) anubis (Hemprich and Ehrenberg, 1833) |
| 哈萨克斯坦亚种 V. v. ochroxantha | 谢尔盖·奥格涅夫, 1926 | 七河地区,阿克赛钦,俄罗斯东部的突厥斯坦,吉尔吉斯斯坦 | ||
| 巴勒斯坦亚种 V. v. palaestina
|
奥德菲尔德·托马斯, 1920 | 以色列雅法附近的拉姆拉 | ||
| 韩国亚种 V. v. peculiosa | 岸田久吉, 1924 | 韩国和中国东北 | kiyomassai (Kishida and Mori, 1929) | |
| 白足亚种 V. v. pusilla
|
爱德华·布莱思, 1854 | 比阿富汗亚种(V. v. griffithii)略小。[33]:123。外观与孟加拉狐(V. bengalensis)类似,但可从长尾和后脚区分。[33]:129 | 巴基斯坦旁遮普省盐岭地区 | leucopus (Blyth, 1854) persicus (Blanford, 1875) |
| 北部平原亚种 V. v. regalis
|
克林顿·哈特·梅里厄姆, 1900 | 北美体型最大的赤狐亚种,耳朵宽大,尾巴长。毛发金黄色,尾巴黑色。[30] | 美国明尼苏达州舍本县麋鹿河附近 | |
| 新斯科舍省亚种 V. v. rubricosa | 欧南·班, 1898 | 体型较大,尾巴宽且长,牙齿和吻突比北美亚种(V. v. fulvus)更长,毛皮颜色深。[30] | 加拿大新斯科舍省迪格比县 | bangsi (Merriam, 1900) deletrix (Bangs, 1898) |
| 萨哈林亚种 V. v. schrencki
|
岸田久吉, 1924 | 俄罗斯库页岛,日本北海道 | ||
| 伊比利亚亚种 V. v. silacea
|
格里特·史密斯·米勒, 1907 | 体型与斯堪的纳维亚亚种(V. v. vulpes)相似,但牙齿更小,前磨牙间隔更宽。毛皮不带红黄色调。身体后半部灰白色,尾巴灰色。[36] | 伊比利亚半岛 | |
| 库页岛亚种 V. v. splendidissima | 岸田久吉, 1924 | 俄罗斯库页岛中部和北部 | ||
| 草原亚种 V. v. stepensis | Brauner, 1914 | 该亚种体型比欧洲亚种(V. v. crucigera)略小,毛发更短、粗糙,颜色更鲜艳。采集自克里米亚山脉的标本具有更明亮、更蓬松和更密集的毛发。[6] | 乌克兰赫尔松附近的草原 | krymeamontana (Brauner, 1914) crymensis (Brauner, 1914) |
| 托博尔斯克亚种 V. v. tobolica | 谢尔盖·奥格涅夫, 1926 | 体型较大,毛发颜色呈铁锈黄或暗红色,腹部为黑色。被毛长且蓬松。[6] | 俄罗斯托博尔斯克和萨列哈尔德 | |
| 华北亚种 V. v. tschiliensis
|
保罗·马奇, 1907 | 体型略大于华南亚种(V. v. hoole) , 与斯堪的纳维亚亚种(V. v. vulpes)类似。[37] | 中国东北部,北京附近 | huli (Sowerby, 1923) |










.jpg.webp)


_fur_skin.jpg.webp)
.jpg.webp)
.jpg.webp)


.jpg.webp)





_Tal_Chappar_Rajasthan_India_14.02.2013.jpg.webp)
.jpg.webp)


_fur_skin.jpg.webp)
分布与栖息地

赤狐的分布范围十分广泛,覆盖了接近7000万平方千米的土地,向北可延伸到北极圈。整个欧洲都有赤狐的足迹,此外它们还分布于非洲撒哈拉以北地区,除东南亚以外的亚洲,除美国西南部和墨西哥外的北美洲。格陵兰岛、冰岛、北极群岛,西伯利亚最北端和极端沙漠中没有赤狐的活动。[8] 新西兰没有赤狐的分布,并且根据新西兰《1996有害物质和外来生物法案》,赤狐被禁止引入该国。[38]
澳大利亚
根据2012年的估计,在澳大利亚有超过720万只赤狐[39],活动范围遍布澳洲各处。[40] 赤狐是在1830-1840年间从英国引入澳大利亚的,最初目的是为了在当地进行英国传统的猎狐活动。在澳大利亚南部岛屿塔斯马尼亚州没有永久定居的赤狐,这或许是因为它们在与袋獾的竞争中处于劣势。[41] 不过在澳洲大陆,赤狐已成为当地的顶级掠食者。在澳洲野犬生活区域,赤狐的活动较少,不过它也借助挖掘洞穴的技能取得了与野猫和野狗的生态位分化。因此,赤狐成为了澳洲大陆最具破坏性的入侵物种之一。
已知赤狐至少造成了多个澳大利亚本土物种的减少或灭绝,尤其是鼠袋鼠科动物,例如荒漠袋鼠。[42] 赤狐在澳洲南部的散布与兔子在澳洲的散布相似,并造成了多种中型地栖哺乳动物的衰退,这些动物包括:岩袋鼠、勒氏短鼻鼠、赤褐袋鼠属、兔耳袋狸属、袋食蚁兽、尖尾兔袋鼠、短尾矮袋鼠。[43] 上述大部分受威胁物种如今都只生活于没有赤狐活动的小岛。澳大利亚曾发起赤狐根除项目,但由于赤狐偏爱穴居和夜间活动,此类项目很难取得有效成果。因此,目前对此问题的解决方法主要集中在管理族群数量,包括引入国家赏金机制。[44] 根据塔斯马尼亚政府的报告,在1999-2000年左右,赤狐被意外引入了先前没有活动的岛屿,对当地原生动物(例如盖氏袋鼠)造成了威胁。因此,塔斯马尼亚州自然资源与环境部组织了当地的赤狐根除计划。[45]
形态特征

外观
赤狐身体较长,四肢相对较短。它们的尾长超过体长一半,大致相当于头部加身体长度的七成,形态蓬松,站立时尾巴可以触地。[6][47] 赤狐的瞳孔为垂直的椭圆形[6],眼部有瞬膜,但只在闭眼时可活动。[7] 赤狐前脚有五个脚趾,后脚则只有四个脚趾,且后脚没有上爪。[7] 赤狐十分敏捷,可跳过两米高的围栏,也会游泳。[48] 通常雌赤狐有4对乳头[6],不过也常有雌赤狐有7个、9个或10个乳头。[7] 雄性赤狐的睾丸比北极狐小。[6]
赤狐的头部窄长,颅骨较小。它们的犬齿较长,头部的两性异形特征比沙狐更明显:雌性赤狐的头骨小于雄性,鼻部区域更宽,腭更硬,犬牙更长。[6] 与狗相比,赤狐的吻突更窄,前臼齿间隙更宽,犬齿细长,轮廓凹凸不平。[7]
体型
赤狐是体型最大的狐属生物[49],不过它的体重要比体型类似的犬属轻。例如赤狐的四肢骨比相似体型的狗的四肢骨轻30%左右。[50] 根据个体、性别、年龄和地理差异,赤狐的体型各不相同。平均而言,成年赤狐肩高35-55.5cm,体长45-90cm,尾长30-55.5cm。耳朵长度7.7-12.5cm,后脚12-18.5cm。体重在2.2-14kg之间,雌性赤狐通常比雄性轻15-20%。[51][52] 成年赤狐头骨长度129-167mm,雌性赤狐128-159mm。[6] 前脚掌长度约60mm,宽度约45mm,后脚掌长度约55mm,宽度约38mm。赤狐的跑步速度一般为6-13km/h,最大奔跑速度约为50km/h。赤狐正常行走时的步幅约为25-35cm。[40] 生活于北美的赤狐体型一般较小,体长相对较长,两性异形更显著。生活于英国的赤狐体型大,体长短,而生活于欧洲大陆的赤狐则比较接近赤狐族群的平均体型。[49] 英国有记录体型最大的赤狐达17.2kg,1.4m长,这只赤狐是雄性,在2012年苏格兰的阿伯丁郡被猎杀。[53]
毛皮

赤狐的冬毛通常致密、柔软、光滑,并相对较长。栖息地偏北的赤狐毛发更长,密而蓬松,而南方赤狐的毛发则更短、更稀疏。[6] 在生活于北部的赤狐中,北美亚种的绒毛比欧洲亚种更柔软。[12] “热窗”部位,例如头和下肢部位毛发终年密而短,其它部位毛发则随季节变化。赤狐的血管舒张与收缩控制了这些部位的热量散出。[54]
赤狐毛皮颜色可分为三类:红色、银/黑色和杂色。[55] 典型的红色毛皮呈现出混有黄色调的铁锈红。沿脊柱有一条较浅的棕色-红色-栗色条纹。另外两条纹路穿过肩胛骨,与脊椎条纹十字交错。后背部通常混有银色,侧边颜色比背部浅,而下巴、下唇、喉咙和前胸则都为白色。下身剩余部分呈黑色、棕色或红色。[6] 哺乳期的雌赤狐肚子可能变为砖红色。[7] 赤狐四肢上部为红锈色,脚掌黑色。面部和上颈前部是明亮的棕锈红色,上唇白色,耳背黑色或棕红色,耳内表面白色。尾巴上部是比背部和侧腹部更浅的棕红色,下部浅灰。黑色的尾上腺通常位于尾巴基部。尾尖为白色。[6]
颜色变体

赤狐毛皮的非典型色彩通常是向黑化的过渡[6],这种现象大多发生在寒冷区域。[9]
| 颜色 | 图片 | 描述 |
|---|---|---|
| 红色 | .jpg.webp) |
最典型的赤狐毛皮颜色 |
| 灰色 | 脊柱与后部呈棕色或灰色,护毛上有浅黄色的条纹。背部的交错条纹为棕色、锈棕色或红棕色。四肢棕色。[6] | |
| 交错 | _Norway_%2526_Canada.jpg.webp) |
毛发的颜色更深,后背部为深棕色或深灰色,护毛上镶有不同程度的银色。背部交错呈黑色或棕色,有时还带有银色。头和脚是棕色的。[6] |
| 棕黑色 | 欧亚赤狐的黑化形态,基本为棕黑色,参杂有不同程度的银色。红色毛发非常少或几乎没有。[6] | |
| 银色 |  |
北美赤狐的黑化形态,有时经由皮毛贸易带入旧世界。特点是颜色纯黑,通常还混有不同程度(25-100%)的银色。[6] |
| 白金色 | _fur_skin.jpg.webp) |
相比银色变体,白金色变体更浅,几乎纯银白,带有蓝色调。[12]:251 |
| 琥珀色 | _fur_skin.jpg.webp) |
|
| 参孙变体 |  |
没有护毛,外观类似羊毛。 [12]:230 |
生活习性
_-British_Wildlife_Centre-8.jpg.webp)


领地标记
赤狐要么在特定范围内定居,要么无居所四处游荡。[40] 它们使用尿液标记领地。[58][59] 排尿时,雄性赤狐抬起一只后脚,排尿方向朝自己身前;雌性赤狐则是蹲下排尿,尿液落后腿之间的土地上。[60] 此外,尿液也被用于标记用来储存食物的空区域以供备用。[40][61][62] 赤狐使用多达12种不同的排尿姿势,以精确的控制气味标记。[63] 生活于同一家庭的赤狐共享一片领地。在环境适宜,食物充足的区域,可能会有一些从属赤狐,它们数量一般为一到两只,有时可多达八只。从属赤狐可能是先前团体的优势阶,但现在已被团体中的年轻成员取代,如今负责照料群体中的幼崽。也有假说提出,从属赤狐的存在也可能与照料幼崽无关,可能是由于食物过剩的结果。从属赤狐不担任繁殖角色,它们负责守卫、玩耍、梳理毛发、照顾幼崽[7],这是亲属选择的案例之一。假如刚成年的赤狐有机会取得新的领地,它们就会离开目前的家庭,否则它们仍会与父母生活在一起,并推迟自己的繁殖期。[40]
繁殖发育
.jpg.webp)
赤狐的繁殖期在每年春季。在发情期前两月(通常是上一年的12月),雌赤狐的繁殖器官开始改变形态与尺寸。进入发情期后,雌赤狐的子宫角扩大为原来2倍大小,卵巢也扩大为原来的1.5-2倍。在上一年8月至9月,雄赤狐就开始产生精子,睾丸在12月至来年2月达到最大重量。[6] 雌赤狐发情期持续两周[7],在此期间,雌雄赤狐通常会在洞穴交配数日。交尾时,雄性赤狐的龟头球会增大[64],交配时间可能持续一个多小时。[7] 怀孕期持续49-58天。[6] 尽管赤狐是单配偶制的[65],但DNA证据显示,赤狐中常有一夫多妻、近亲性交、单窝幼崽父母各不相同的情况。[7] 团体中的从属赤狐也可能会怀孕,但它们通常无法顺利产仔,即使成功产下幼崽,它们的后代也可能会被优势阶雌赤狐或其它从属赤狐杀害。[7]

通常赤狐一胎可产4-6个幼崽,偶尔也记录到一胎多达13个幼崽。[6] 在赤狐死亡率较高的区域,幼崽出生数也较多。[40] 刚出生的赤狐不具备视觉与听觉,也没有牙齿,毛发是蓬松的暗棕色。新生赤狐体重在56-110g,体长约14.5cm,尾长约7.5cm。小赤狐头大、腿短、胸宽。[6] 赤狐母亲会持续照料小赤狐2-3周,直到它们能够进行体温调节。在这段时间里,产仔的雌赤狐由雄性赤狐或其它未产仔的雌赤狐负责喂食。[7] 雌赤狐会努力保护幼崽,曾观察到它们能够击退猎犬。[25]:21-22 假如赤狐母亲在小赤狐独立前死去,赤狐父亲会接过照料幼崽的责任。[25]:13 小赤狐的眼睛在13-15天后张开,在此期间它们的耳道也会开启,上牙开始长出,下牙的生长晚3-4天。[6] 小赤狐的眼睛最初是蓝色的,在4-5周后转变为琥珀色。毛发颜色的变化始于3周左右,此时眼睛周围的毛发开始变深。一个月时,赤狐的面部出现红色与白色毛发,耳朵开始耸立,吻突变长。[7] 在3-4周龄时,赤狐开始在巢外活动,并食用父母提供的固状食物。赤狐的哺乳期持续6-7周。[6] 8周后,赤狐绒状的毛发开始被光亮的护毛覆盖。[7] 到3-4个月时,小赤狐的腿已变长,胸窄且肌肉发达。它们在6-7个月时达到与成年赤狐相当的体型。[6] 有些品种的赤狐可在9-10个月时性成熟,因此可在1岁时产下首胎。[6] 圈养环境中的赤狐寿命可达15年,但在野外它们通常只能存活5年。[66]
挖掘洞穴
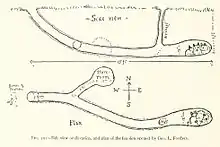
在非繁殖时期,赤狐一般在植被茂密的开阔地带活动,但有时也会挖掘并躲入洞穴以躲避恶劣天气。[7] 洞穴通常位于山坡、沟壑、断崖、河岸峭壁、岩石缝隙、水沟等位置。赤狐偏好在排水良好的土壤中挖掘洞穴,在树根之间挖掘的洞穴可维持数十年,而在草原上挖掘的洞穴则只能维持几年。[6] 赤狐可能会在毛囊虫病爆发时废弃现有洞穴,这或许是为了阻止疾病的传播。[7] 在欧亚大陆的沙漠地带,赤狐可能会利用狼、豪猪或是沙鼠遗留的洞穴。相比獾、土拨鼠以及其它狐属成员,赤狐的洞穴并不复杂。赤狐的洞穴可分为长期洞穴与临时洞穴,后者可能仅是一个用于临时躲藏的小通道或洞窟。从洞穴入口向下的坡度约为40-45°,底部的巢穴空间宽阔,并延伸出数条分支隧道。洞穴宽度在0.5-2.5米之间,一般不与地下水相连。洞穴长度一般在5-7米,最长可达17米。春天时,赤狐会清理巢穴内多余的土壤,清理速度很快,它们先用前脚挖土,然后用后脚将挖出的土壤踢到距离洞穴2米外的位置。当小赤狐出生后,挖出的土堆会被踏平,作为它们玩耍和接受食物的场所。[6] 赤狐可能会与土拨鼠[9]或獾分享洞穴。[6] 赤狐卫生习惯不如獾那样讲究,獾会细致的清理巢穴,并有专门的“厕所”,而赤狐则随意将食物碎屑丢在洞穴周围。[25]:15-17 圈养环境中的赤狐睡眠时间通常为9.8小时/天。[67]
交流方式
肢体语言
_WOB.JPG.webp)
赤狐的肢体语言由耳朵、尾巴的活动以及不同的身体姿态组成。肢体语言传达的意义可分为进攻/主导和畏惧/臣服两类。有些姿态可能共同包含这两类。[40]:42-43 嗅探时,赤狐的耳朵会旋转并抽动。玩耍时,它们的耳朵竖起,用后脚站立。雄赤狐在向雌性求爱时,或成功赶走入侵者后,它们的耳朵会向外翻转,尾巴抬起呈水平状,尾尖朝上。感到害怕时,赤狐会表现出臣服的姿势,此时它们弓起背部,身体弯曲,四肢下蹲,尾巴前后拍打,耳尖向后贴在头上。假如只是为了表达臣服,姿势与上述类似,但不会有弓起背部和弯曲身体的动作。当两只势均力敌的赤狐争夺食物,它们会相互靠近,以侧身推挤对方,姿态共同混有进攻和畏惧的成分,此时它们拍打尾巴、弓起背部,但不下蹲,耳朵也不朝后贴在头上。[68] 当赤狐发起自信的攻击时,它会径直冲向对方,而不是从侧边擦过,此时它的尾巴抬起,耳朵向外翻转。[40] 在这种争斗中,赤狐会将前腿抵对方的身上,张嘴相互威胁。一般情况下,这类争斗发生在青少年赤狐或同性别成年赤狐之间。[7]
叫声
赤狐的发声范围很广,不同叫声可跨越五个八度,这些叫声被分为不同等级。[40]: 28 最近分析识别出幼年和成年赤狐的12种不同叫声。[7] 大部分叫声可被归为“联络”和“交互”两类。前者由交流双方相隔距离而变化,后者则由进攻性的程度而变化。[40]: 28
- 联络叫声:最常见的联络叫声是由三到五个“wow wow wow”音节构成的。这个叫声常出现在在12月至2月间,并有可能与灰林鸮的叫声混淆。“wow wow wow”叫声随个体不同而有所差异;曾有记录圈养中的赤狐只会回应伴侣的,而不会回应其它陌生赤狐的录制叫声。幼年赤狐大约在19天时可发出“wow wow wow”的叫声,目的是渴望得到关注。当赤狐相互靠近,它们会发出类似于鸡叫的三音节叫声。成年赤狐会用粗糙的喘息声来向自己的幼崽打招呼。[40]: 28
- 交互叫声:赤狐使用高亢的呜咽声相互招呼,尤其是从属赤狐。当从属赤狐被另一优势阶赤狐靠近时,它们会发出类似警报器的尖叫声。当赤狐之间相互攻击时,它们会发出一种类似棘轮的喉音,这种叫声被称为“gekkering”。Gekkering主要出现于求偶季节,来自于相互竞争的雄赤狐或拒绝求爱的雌赤狐。在驯养和被驯化的赤狐中曾被观察到会发出类似笑声的声音,这种叫声被认为是为了和它们的人类饲养者沟通。[69][70]
此外,还有一种无法被归为上述两类的叫声,这个叫声是单音节的长呼号“waaaaah”。这个叫声常见于繁殖季节,被认为是雌赤狐呼唤雄性所用。感到威胁时,赤狐也会发出单音节叫声。距离较近时,这个叫声类似低沉的咳嗽,距离较远时,声音更加尖锐。哺育中的赤狐幼崽会发出呜咽声,当需求得不到满足时,这些声音会变得十分吵闹。[40]: 28
生态学
食物

赤狐是食肉动物,食物来源十分多样。前苏联科学家曾记录赤狐食用300种不同动物以及数十种不同植物。[6] 赤狐主要捕食小型啮齿类,例如田鼠、小鼠、地松鼠、仓鼠、沙鼠[6]、土拨鼠、囊鼠、白足鼠等。[9] 其次,赤狐还捕食鸟类(主要是燕雀、野鸡、雁鸭)、兔子、豪猪、浣熊、负鼠、爬行动物、昆虫、无脊椎动物和水产。[6][9] 在极少数情况下,赤狐会试图猎食有蹄类幼崽。[6] 赤狐的攻击目标一般是3.5千克体重以下的动物,每天摄食大约500克。[49] 赤狐有时也吃植物,在部分地区,秋天时赤狐可完全依赖水果存活。赤狐常食用的水果包括蓝莓、树莓、黑莓、樱桃、柿子、桑葚、苹果、李子、葡萄和橡子。此外赤狐也偶尔食用禾本科、莎草科植物,以及块茎与块根。[9]
虽然人们普遍认为嗅觉在赤狐的捕猎过程中占据主导地位[71],但两项实验发现,在视觉、嗅觉和听觉中,赤狐[72]和郊狼在狩猎时主要用到的是视觉。[73][74]
赤狐偏好在深夜以及日出前的清晨狩猎。[6] 尽管它们一般单独行动,但在资源丰富的地区仍会有许多赤狐聚集。[66] 在捕食类似老鼠的动物时,赤狐会先依据声音定位,然后跃起,在半空中使用尾巴调整方向,落在5米内的位置。[75] 傍晚和夜间赤狐一般只寻找腐肉。[6] 它们对食物有极强的占有欲,甚至拒绝将其分享给当前领地的优势阶赤狐。[40]:58 赤狐有时会做出过捕行为;曾有记录在繁殖季节,四只赤狐在天黑风大、飞行状况不好的情况下共猎杀了约200只红嘴鸥。这种行为可能会对家禽数量造成潜在打击。[7][40]:164 赤狐似乎不喜欢鼹鼠的味道,但它们仍会活捉鼹鼠给自己的幼崽玩耍。[40]:41
一项2008-2010年在捷克和德国进行的研究发现,赤狐在茂密植被或雪地中的成功狩猎似乎是依靠地球磁场的帮助。[40]:41
竞争者

赤狐通常在与其它狐属的竞争中占据优势。北极狐因不敌赤狐而往北迁徙,北部地区食物稀少,赤狐难以获得足够的食物以维持较大体型。赤狐栖息地的北部边界与可获取的食物量相关,而北极狐栖息地的南部边界与赤狐的存在相关。由于1830-1930年蓬勃发展的皮草贸易,赤狐和北极狐被引入了从阿留申群岛至亚历山大群岛之间的几乎每个岛屿。赤狐可以轻易地取代北极狐,1866年曾有记录一只雄赤狐猎杀了一个小岛上所有的北极狐。[40] 在共域地区,北极狐可能会放弃捕食赤狐最爱的田鼠,转而捕食旅鼠和水生动物。只要有机会,赤狐和北极狐会尝试杀死对方的幼崽。[6] 沙狐也是赤狐的竞争者之一,它们几乎全年捕食相同的猎物。相比沙狐,赤狐更强壮,更适应在超过10cm的雪地捕食,并能更加有效率的捕捉大中型啮齿类。沙狐似乎只在半沙漠和草原地带能够胜过赤狐。[6] 以色列的阿富汗狐为了躲避与赤狐的竞争而居住在山崖地带,并积极避免在开阔平地上与赤狐照面。[40]:84-85 敏狐和草原狐也受赤狐的压制。为避免与赤狐发生竞争,敏狐一般居住在更加干旱的环境,不过随着人类对赤狐的引入,许多先前生活着敏狐的地区如今都已被赤狐占据。赤狐会捕食它们,并占据它们的巢穴。[9] 灰狐与其它狐狸不同,它们在于赤狐的竞争中处于优势。历史上少有灰狐与赤狐碰面的记录,因为灰狐偏好密林或半干旱栖息地,而赤狐则主要在开阔、湿度中等的区域活动。不过由于森林砍伐,两个物种间的交互变得越加频繁。[9]
狼可能会在于赤狐争夺动物尸体时杀掉并食用赤狐。[6][76] 在北美某些地区,当赤狐与郊狼共域时,赤狐会倾向于在郊狼领地范围外定居,这或许是它们为了躲避郊狼而作出的刻意选择。赤狐与郊狼相遇时的行为各不相同,可能主动接触,也可能相忽视。大多数进攻行为是由郊狼发起的,赤狐主动攻击郊狼的记录十分少见,除非它们的幼崽受到威胁。赤狐和郊狼也曾被观察到共同进食。[77] 在以色列,赤狐与亚洲胡狼分享栖息地。在领地重合区,赤狐与胡狼会因共同食物来源而发生竞争。在其领地内,赤狐会无视胡狼的气味或踪迹,并避免与胡狼发生近距离身体接触。在胡狼族群充裕的地区,赤狐的数量会显著下降,这显然是由竞争排除原则造成的。[78]
.jpg.webp)

赤狐在与貉的竞争中有占有优势,它们有时会杀死貉的幼崽,或咬死成年貉。有记录赤狐进入貉的洞穴并将它们杀死。赤狐与貉都以类似老鼠的小啮齿类为食,这种竞争春季时尤为激烈,因为此时食物来源较少。在鞑靼斯坦共和国的调查显示,在56只死亡的貉中,有11.1%是由赤狐造成的;俄罗斯西北部的另一项调查显示,在186只死亡的貉中,有14.3%是由赤狐造成的。[6]
赤狐也可能会杀死鼬科动物,例如伶鼬、石貂、松貂、白鼬、黄鼬、欧洲鼬以及年幼的紫貂。欧洲狗獾可能与赤狐生活于同一洞穴,它们居住在洞穴中的孤立部分。[6] 欧洲狗獾和赤狐可能是互利共生的关系:赤狐为欧洲狗獾提供食物,而欧洲狗獾帮助赤狐维持洞穴清洁。[25]:15 但也有观察到欧洲狗獾将雌赤狐从洞穴中驱离,并杀死赤狐幼崽,不过并没有将它们吃掉。[79] 貂熊可能会趁赤狐睡觉或食用腐肉时将它们杀死。[6]:546 赤狐也会杀死貂熊幼崽。[80]
赤狐可能会与条纹鬣狗争夺大型动物尸体。赤狐会将未咬开的尸体让给条纹鬣狗,因为它们的牙齿较难撕开坚硬的肉体,而条纹鬣狗则可以轻易做到这一点。条纹鬣狗取食时,赤狐会上前骚扰,并借助较小的身躯、敏捷的动作躲开条纹鬣狗的回击。即使是在没有食物的情况下,赤狐似乎也会这样故意折磨条纹鬣狗。不过有时候赤狐会失手被条纹鬣狗杀死。[40]: 77–79 条纹鬣狗巢穴中常有赤狐的尸体,有时条纹鬣狗也会偷走掉入陷阱的赤狐。[6]
在欧亚大陆,赤狐可能会被豹、狞猫和欧亚猞猁捕食。欧亚猞猁会在厚雪地上追逐赤狐,借助更长的腿和更大的脚掌,它们能轻易追上赤狐,在超过一米厚的雪地上则更为轻松。[6] 在俄罗斯 Velikoluki 地区,赤狐极少出现在欧亚猞猁的领地内。[6] 研究者认为欧亚猞猁对赤狐造成的威胁远小于狼的威胁。[6] 生活于北美的赤狐面临的威胁主要是美洲狮、加拿大猞猁和短尾貓。[47] 有时,雕鸮会捕食幼年赤狐,极少情况下也会捕食成年赤狐。[81][82] 已知金雕、楔尾雕、白尾海雕和虎头海雕会捕食成年赤狐。[83][84][85][86]
疾病

在欧洲,赤狐是主要的狂犬病携带者。生活在伦敦的赤狐常患有关节炎,患病位置主要在脊椎。[7] 赤狐可能感染钩端螺旋体病和兔热病,不过后者对赤狐造成的影响很小。它们还可能患上李氏菌病和肠道旋体病,并作为载体传播丹毒、布鲁氏菌病和蜱虫脑炎。在俄罗斯萨尔特兰湖附近的赤狐族群中曾记录到一种神秘的致命传染病,但原因未确定,可能是严重的脑脊髓炎,最初在人工饲养的银狐观察到这种疾病。已知赤狐可能会被鼠疫杆菌感染。[6]
赤狐一般不受跳蚤的侵扰。赤狐可能会在捕食某些猎物时染上欧洲兔跳蚤,在旅行时染上刺猬跳蚤。可在赤狐身上生活的跳蚤包括人蚤、狗蚤和獾蚤。赤狐身上的蜱虫也比较常见,例如篦子硬蜱和六角硬蜱,它们较多出现在哺育期的雌赤狐及赤狐幼崽身上。虱毛目中的 Trichodectes vulpis 是专门寄生于赤狐的品种,但较为少见。螨虫中的疥螨是造成赤狐罹患毛囊蟲病的主要原因。毛囊蟲病可造成明显的毛发脱落,脱落始于尾部和后腿,然后向身体其他部位扩展。在毛囊蟲病后期,赤狐会失去大部分毛发,体重降低50%,并啃咬受感染的四肢。毛囊蟲病疫情蔓延时,染病的赤狐大约会在4个月后死去。影响赤狐的其它内寄生虫还包括:毛囊蠕形螨、背肛螨(Notoderes)、耳螨(常发现于耳道)、锯齿舌形虫(常感染鼻腔)和皮癣菌病。[6]
在毛皮牧场的赤狐中曾发现多达60种寄生虫,其中有20种是已知存在野外的。已知球虫亚纲下的联同形球虫属和艾美球虫属能够感染赤狐。[6] 赤狐中常见的线虫包括犬蛔虫、狭头钩虫、Capillaria aerophila和犬线虫(Crenosoma vulpis),后两种线虫主要感染赤狐的肺部。[87] 犬膀胱虫感染赤狐的膀胱。旋毛虫对赤狐的影响不大。赤狐中常见的真绦虫包括旋绦虫和豌豆状绦虫。其它还包括单包绦虫和多包绦虫。有11种吸虫可感染赤狐[7],包括结合次睾吸虫。[88]
与人类的关系
文化形象


赤狐在不同民间文化中都占据有重要的地位。希腊神话中曾描绘了一只身形巨大,永远无法被捕捉的透墨索斯恶狐。[89][90]
在凯尔特神话中,赤狐是一种重要的动物。例如在英国科茨沃尔德,当地人相信巫师会化身为赤狐偷取邻居家的黄油。[91] 在后来的欧洲民俗中,列那狐的形象象征了狡诈与欺骗。列那狐最初是公元1150年的诗歌《Ysengrimus》里的一个次要角色,而后在1175年被皮埃尔·圣克劳德编入了小说《列那狐的故事》,后来又出现在杰弗里·乔叟的《修女牧师故事集》中。许多有关列那狐的冒险故事都取材于赤狐在野外的真实习性,例如列那狐与狼为敌,喜爱黑莓与葡萄。[40]:32-33
中国民间传说中也有以赤狐为原型的“狐狸精”,在一些故事中狐狸精有九条尾巴,韩国将其称为“Kumiho”。[92] 在日本神话中有被称为“kitsune”的超自然生物,它是一种拥有魔法能力的狐狸,并且随年龄增长变得越来越聪明。在 kitsune 拥有的魔法中,最重要的是它幻化为人类的能力。在一些故事里,kitsune 使用这个能力戏耍他人,另一些故事则把 kitsune 描绘为一个忠实的守卫、朋友、爱人或妻子。[93] 在阿拉伯民俗文化中,赤狐被认为是一种怯懦、软弱、狡猾、善欺骗的动物,人们认为赤狐会鼓起肚子侧卧在地上装死,等待不知情的猎物靠近。[35] 《圣经》借狐狸狡猾的习性讽刺那些虚假的先知[94],以及希律·安提帕斯的虚伪。[95][96]
美洲原住民神话中也有狡猾的狐狸形象出现,它常被描述为是郊狼的伙伴。故事中的赤狐是个不忠实的伙伴,它常偷吃郊狼的食物。在阿霍马维创世神话中,赤狐与郊狼携手创造了天地,并在人类到来前离开了世界。尤罗克人相信赤狐在愤怒之中从天上取下太阳,将其安放在山顶,使地面燃烧出一个巨大的坑洞。因纽特神话故事讲述了一只化为女人的赤狐诱骗一个猎人与她结婚,后来因受到猎人的冒犯而化为原形离去。梅诺米尼人的故事讲述了赤狐如何作为狼的不忠诚伙伴。[97]
狩猎活动

最早猎杀赤狐的历史记录可追溯到公元前4世纪;亚历山大大帝曾狩猎赤狐,一个公元前350年的印章描绘了一位波斯猎人用矛刺杀赤狐的场景。古希腊历史学家色诺芬曾鼓励人们猎杀赤狐,他认为这可作为一种教育手段,并且他还认为赤狐是一种有害生物,因为它们分散了猎狗的注意力。罗马人在公元80年左右就开始猎杀赤狐。在欧洲黑暗时代,赤狐被认为是次要的狩猎目标,但后来逐渐受到重视。克努特大帝将赤狐重新归类为“值得追猎的目标”,仅次于“最有价值的狩猎目标”。赤狐逐渐被作为逐猎目标而非有害生物猎杀,以至于到了12世纪,爱德华一世拥有一队皇家猎狐犬和一个专门的猎狐人。到了文艺复兴时期,猎狐已经成了贵族们的一项传统运动。尤其在英国内战后,鹿的数量大幅下降,猎狐活动变得越来越受欢迎。首个猎狐俱乐部“查尔顿猎狐俱乐部”成立于1737年,猎狐活动在18世纪达到了最高峰。[40]:21 尽管北美也生活有本土赤狐,但在1730年代,弗吉尼亚和马里兰州富裕的烟草农场主仍从欧洲大陆引入了赤狐,用作运动消遣。 这些人认为欧洲赤狐比美洲赤狐更适合猎狐运动。[98]
灰狐更有趣,而赤狐更刺激。灰狐跑的没那没快,但经常绕着自己巢穴奔跑,一圈大约六到八英里。而老赤狐,我叫它老赤狐不是因为年龄,而是对它能力的赞赏,它可以带着猎狗跑一整天,直到天黑,不断奔跑30英里后甩开它们离去。所以虽然捕获灰狐可以得到人们的夸奖,但只有捕获“老赤狐”才能成为他们的谈资。[98]
如今,赤狐仍被许多人当做有害生物猎杀,人类是造成赤狐死亡的最主要因素。每年赤狐捕杀量如下:英国 21,500–25,000 (2000); 德国 600,000 (2000–2001); 澳大利亚 58,000 (2000–2001); 瑞典 58,000 (1999–2000); 芬兰 56,000 (2000–2001); 丹麦 50,000 (1976–1977); 瑞士 34,832 (2001); 挪威 17,000 (2000–2001); 加拿大萨斯喀彻温省 2,000 (2000–2001); 加拿大新斯科舍省 491 (2000–2001); 美国明尼苏达州 4,000–8,000 (2002–2009年均); 美国新墨西哥州 69 (1999–2000)。[49]
毛皮贸易


赤狐是皮草贸易中最重要的动物之一。它们的毛皮被用在衣饰、围巾、手笼、夹克和皮草的制作中,主要用作布料服装的衣饰,或是用于制作整件毛皮大衣(包括女宽罩衫)。[12]:229-230 银狐的毛皮广泛用来制作披肩[12]:246,混色毛皮则主要用来制作围巾,偶尔作为衣饰。[12]:252 以赤狐毛皮为材料的围巾比其它动物毛皮围巾的总量还多,但是仍比不上赤狐毛皮在衣饰方面的普遍程度。[12]:229-230 毛皮颜色的受欢迎程度依次为银色、混色和红色。[25]:207 在20世纪初,每年有超过1000张赤狐毛皮从美国出口至英国,而从俄罗斯和德国出口的赤狐毛皮更是高达500,000张。[25]:6 根据1985-1986年的数据,全球赤狐毛皮交易量为1,543,995张。赤狐占了美国野外捕获毛皮的45%,总额达5000万美元。[49] 毛皮价格在持续上涨,2012年赤狐毛皮在北美的批发拍卖价为39美元/张,2013年则涨到了65.78美元/张。[99]
北美赤狐,尤其是生活于阿拉斯加北部的赤狐亚种的毛皮价值最高,因为它们的毛皮质地光滑,人们穿上其制作的皮草后仍可活动自如。相比之下,居住在阿拉斯加南部海岸以及阿留申群岛的赤狐亚种的皮毛就粗糙许多,它们的毛皮价格从未超过北部同胞的1/3。[12]:231 相比北美亚种,欧洲赤狐亚种的毛皮更加粗糙。其中唯一的例外是生活在北欧及俄罗斯远东地区的赤狐亚种,但它们的毛皮光滑程度仍不如北美赤狐。[12]:235
攻击家畜


赤狐偶尔会捕食羊羔,它们通常会选择孱弱目标,但也并不总是如此。品种体型小的羔羊(例如苏格兰黑面羊)较容易受到赤狐攻击。多只小羔羊比单只更容易受攻击,因为母羊无法同时保护到它们。小体型高地与大体型低地公羊的杂交后代更容易受赤狐攻击,因为这些后代体型较大,母羊要花费更多精力照料它们,从而暴露出更多赤狐可利用的机会。首次生产的母羊的后代更容易受赤狐攻击,而经验老道的母羊则离自己孩子更近,保护更周全。[40]166-167
如果饲养的家兔和豚鼠被允许在花园或户外自由活动,它们有可能会被赤狐捕食。在城市游荡的赤狐可能常与野猫照面,并更跟随野猫搜集食物。在身体对抗中,野猫通常占据上风。在赤狐杀死野猫的案例中,受害者通常是幼年猫咪。不过赤狐偶尔也会攻击野猫,通常不是为了捕食,而是将它们视为食物竞争者。[40]:180-181
家养驯化
.jpg.webp)
野生赤狐一般不适合作为宠物。[100] 许多被遗弃的赤狐幼崽会被善意的人收养,不过雌赤狐似乎一般不会抛弃后代。真正的赤狐孤儿幼崽十分少见,人们收养的其实多为从自己巢穴走丢的小赤狐。[101] 赤狐幼崽几乎需要时刻照看,还未断奶的赤狐每隔四小时就需要喂食牛奶,无论日夜。断奶后的赤狐十分顽皮,可能会破坏其它毛皮物件、家具和电线。[40]:56 尽管年幼的赤狐对人类较为友好,但当它们达到10周龄时会开始对周围除饲养者外的人类感到恐惧。[40]:61 它们仍维持着野外生活的隐蔽本能,并且即使在喂饱的情况下也可能攻击家鸟。[25]:122 尽管对陌生人会感到恐惧,但它们可以和家中的猫狗建立亲密关系。家养的赤狐曾被用于驱赶鸭子。[25]:132-133
毛色黑白的赤狐曾被挑选繁衍,用于生产毛皮,它们被称为“银狐”。20世纪下半叶,俄罗斯动物学家德米特里·别利亚耶夫开始了超过40年的银狐驯化工作。最初,别利亚耶夫保留了银狐后代中对人类恐惧程度较低的个体。随后,别利亚耶夫团队从银狐后代中筛选那些对人类行为作出友善回应的个体进行后续繁衍。随着驯化的进行,银狐的行为和外貌都发生了显著的变化。经过10代有控制的繁衍后,银狐已不再表现出对人类的恐惧,并经常摇着尾巴,舔舐人类饲养者以表示亲近。这些行为变化伴随着外貌的改变,包括更长的毛发、耸拉的耳朵、卷曲的尾巴,这种变化与野狼至家犬的变化十分类似。[102]
分布
赤狐成功的适应了城郊地区的生活,尤其是低密度的郊区。[49] 在过去的二十世纪,赤狐在澳大利亚、欧洲、日本和北美城郊地区建立了稳固的地位。赤狐从1930年代开始进入英国城区生活,并于1940年代进入布里斯托尔和伦敦,随后又扩散到了剑桥和诺维奇。如今,赤狐在爱尔兰都柏林已十分常见。有记录赤狐最早自1930年代生活于澳大利亚墨尔本,在瑞士苏黎世则直到1980年代才有赤狐出现。[103] 赤狐常出现在郊区低密度的私人住宅土地,而工业、商业、租赁土地则少有赤狐的踪影。[49] 在工业、商业、租赁土地中,赤狐的族群密度较低,因为它们在这些区域更少依靠人类资源生活;赤狐在这些区域的平均活动范围为80-90公顷,相比之下生活于住宅区的赤狐平均活动范围为25-45公顷。[104]
根据2006年的估计,在伦敦生活有大约10,000只赤狐。[105] 相比野外同伴,在城市生活的赤狐可获取的潜在资源更多,竞争者也更少。城市中的赤狐可能会从垃圾桶中搜寻食物,不过它们大部分饮食仍于野外同伴类似。
行为
城郊赤狐主要在清晨和傍晚活动,并在此期间完成大部分猎食工作。日间较难观察到赤狐的踪迹,不过它们偶尔也会被发现在屋顶晒太阳。城郊赤狐的巢穴通常位于城市中较为隐蔽的区域,或是处于城市边缘地带,它们会在夜晚到访巢穴获取一些食物,并睡在这些巢穴中。赤狐是优秀的食物收集者,它们会吃几乎所有的人类食物,有些喜爱赤狐的人会有意的为它们留一些食物。定期投喂食物的行为能够将赤狐吸引至家中,它们会逐渐习惯人类的存在,会允许人类接近它们,甚至与它们一起玩耍。[104]
控制
城郊赤狐可能会对当地居民带来困扰。它们会偷居民养的鸡,翻垃圾桶,损坏园艺设施。大部分对于城郊赤狐的抱怨都集中在一月至二月初赤狐的发情期,以及四月至八月赤狐幼崽的发育期。[104] 英国禁止在城区猎杀赤狐,一个替代方法是设置陷阱诱捕它们。[106] 不过猎杀对于赤狐种群的影响微乎其微,被杀的赤狐很快就会被新生赤狐或是从其它地区迁入的赤狐所取代。另一个更有效的方法是将赤狐从特定区域驱离,驱离方法包括使用杂酚油、柴油或氨水等阻吓剂。清理并阻塞赤狐巢穴也是一个有效的方法,赤狐可能会因此而不再返回该区域。[104]




參考文獻
- . The IUCN Red List of Threatened Species 2008. [20 September 2013].
- Linnæus, Carl. 10th. Holmiæ (Stockholm): Laurentius Salvius. 1758: 40 [2018-05-12]. (原始内容存档于2012-11-08) (拉丁语).
- . Invasive Species Specialist Group. [2021-03-15]. (原始内容存档于2017-03-16).
- Kurtén, Björn (1968). Pleistocene Mammals of Europe. Weidenfeld & Nicolson.
- Kurtén, Björn & Anderson, Elaine. . Columbia University Press. 15 October 1980: 96, 174. ISBN 9780231037334 (英语).
- Heptner, V. G. . Leiden u.a.: Brill. 1998: 115, 341–365, 453–502, 513–562 [8 July 2016]. ISBN 978-1886106819.
- Harris, Stephen & Yalden, Derek. 4th. Southampton: Mammal Society. 2008: 408–422. ISBN 978-0906282656.
- Nasimovich, A. A.; Bannikov, A. G.; Hoffmann, Robert S. . Leiden: Brill. 1989. ISBN 90-04-08873-3. OCLC 17953816.
- Feldhamer, George; Thompson, Bruce & Chapman, Joseph. second. Johns Hopkins University Press. 2003: 516–530. ISBN 9780801874161 (英语).
- . www.wildlifeonline.me.uk. [2022-07-16]. (原始内容存档于2022-06-10) (英语).
- Fedriani, J. M.; Palomares, F. & Delibes, M. . Oecologia. 1999, 121 (1): 138–148. Bibcode:1999Oecol.121..138F. CiteSeerX 10.1.1.587.7215
 . JSTOR 4222449. PMID 28307883. S2CID 39202154. doi:10.1007/s004420050915.
. JSTOR 4222449. PMID 28307883. S2CID 39202154. doi:10.1007/s004420050915. - Bachrach, Max. third. New York: Prentice-Hall. 1953.
- . PETA UK. 2012-04-26 [2022-01-20]. (原始内容存档于2022-01-20) (美国英语).
- Lindblad-Toh, K.; Wade, C. M.; Mikkelsen, T. S.; et al. . Nature. 2005, 438 (7069): 803–819. Bibcode:2005Natur.438..803L. PMID 16341006. doi:10.1038/nature04338 .
- Kurtén, Björn. . Weidenfeld & Nicolson. 1968.
- PaleoDatabase collection No. 35369 (页面存档备份,存于), authorized by Alan Turner, Liverpool John Moores University. Entry by H. O'Regan, 8 December 2003
- Spagnesi & De Marina Marinis 2002,第222頁
- Aubry, Keith B.; Statham, Mark J.; Sacks, Benjamin N.; Perrine, John D.; Wisely, Samantha M. . Molecular Ecology. 2009-06, 18 (12) [2022-07-17]. doi:10.1111/j.1365-294X.2009.04222.x. (原始内容存档于2022-07-17) (英语).
- Statham, Mark J.; Murdoch, James; Janecka, Jan; Aubry, Keith B.; Edwards, Ceiridwen J.; Soulsbury, Carl D.; Berry, Oliver; Wang, Zhenghuan; et al. . Molecular Ecology. 2014, 23 (19): 4813–4830. PMID 25212210. S2CID 25466489. doi:10.1111/mec.12898.
- Statham, Mark J.; Sacks, Benjamin N.; Aubry, Keith B.; Perrine, John D.; Wisely, Samantha M. . Journal of Mammalogy. 2012-02-16, 93 (1). ISSN 0022-2372. doi:10.1644/11-MAMM-A-033.1.
- Sacks, Benjamin N.; Moore, Marcelle; Statham, Mark J.; Wittmer, Heiko U. . Molecular Ecology. 2011-01, 20 (2) [2022-07-17]. doi:10.1111/j.1365-294X.2010.04943.x. (原始内容存档于2022-07-17) (英语).
- Volkmann, Logan A.; Statham, Mark J.; Mooers, Arne Ø. & Sacks, Benjamin N. . Journal of Mammalogy. 2015, 96 (2): 297–307. doi:10.1093/jmammal/gyv007 .
- Wozencraft, W.C. . Wilson, D.E. & Reeder, D.M. (编). (3rd ed.). Baltimore, Maryland: Johns Hopkins University Press. 2005: 532–628. ISBN 978-0-8018-8221-0. LCCN 2005001870. OCLC 62265494. NLC 001238428.
- Castello, Jose, 2018. Canids of the World. Princeton University Press: Princeton, NJ.
- Dale, Thomas Francis. . London, New York, Bombay: Longmans, Green, and Co. 1906 [8 July 2016].
- Teacher, Amber G. F.; Thomas, Jessica A. & Barnes, Ian. . BMC Evolutionary Biology. 2011, 11 (214): 214. PMC 3154186 . PMID 21774815. doi:10.1186/1471-2148-11-214.
- Kutschera, Verena E.; Lecomte, Nicolas; Janke, Axel; Selva, Nuria; Sokolov, Alexander A.; Haun, Timm; Steyer, Katharina; Nowak, Carsten & Hailer, Frank. . BMC Evolutionary Biology. 2013, 13 (114): 114. PMC 3689046 . PMID 23738594. doi:10.1186/1471-2148-13-114.
- Allen, S. H. & Sargeant, A. B. . Journal of Wildlife Management. 1993, 57 (3): 526–533 [2021-03-15]. JSTOR 3809277. doi:10.2307/3809277. (原始内容存档于2020-10-18).
- Preble, Edward Alexander. . . U.S. Government Printing Office. 1908: 217 [2022-12-21]. (原始内容存档于2022-07-26).
- Merriam, Clinton Hart. . Washington Academy of Sciences. 1900: 663–669. ISBN 9780665167928.
- Cook, Joseph A.; MacDonald, Stephen O. . University of Alaska Press. 2010: 162 [2022-12-21]. ISBN 9781602231160. (原始内容存档于2022-07-26).
- Spagnesi & De Marina Marinis 2002,第221頁
- Pocock, Reginald Innes. . 1941 [8 July 2016].
- Allen 1938
- Osborn, Dale J. & Helmy, Ibrahim. . Field Museum of Natural History. 1980: 376, 679 [9 July 2016].
- Miller, Gerrit Smith (1912). Catalogue of the Mammals of Western Europe (Europe Exclusive of Russia) in the Collection of the British Museum, British Museum (Natural History). Department of Zoology.
- Allen 1938,第353頁
- . legislation.govt.nz. [2022-07-19]. (原始内容存档于2022-06-05).
- "Impacts of Feral Animals". Game Council of New South Wales. Archived from the original on 18 April 2012. Retrieved 29 May 2012.
- Macdonald, David W. . Unwin Hyman. 1989. ISBN 0-04-440419-0. OCLC 19776630.
- Bostanci, Adam. . Science. 2005-02-18, 307 (5712) [2022-07-20]. ISSN 0036-8075. doi:10.1126/science.307.5712.1035. (原始内容存档于2022-07-20) (英语).
- Short, Jeff. . Biological Conservation. 1998-12-01, 86 (3) [2022-07-20]. ISSN 0006-3207. doi:10.1016/S0006-3207(98)00026-3. (原始内容存档于2017-09-09) (英语).
- New South Wales National Parks and Wildlife Service. . Hurstville, N.S.W.: NSW National Parks and Wildlife Service. 2001. ISBN 0-7313-6424-4. OCLC 223257960.
- Johnson, Chris. . Cambridge [England]: Cambridge University Press. 2006. ISBN 0-521-84918-7. OCLC 63187432.
- "Latest Physical Evidence of Foxes in Tasmania (页面存档备份,存于)". Department of Primary Industries and Water, Tasmania website. 18 July 2013. Retrieved 20 September 2013.
- . Quaderni di conservazione della natura. 2001: 222 [2022-07-20]. ISSN 1592-2901. OCLC 183253519. (原始内容存档于2022-07-20) (意大利语).
- Larivière, Serge & Pasitschniak-Arts, Maria (1996). Vulpes vulpes (PDF). American Society of Mammalogists. Archived from the original (PDF) on 31 October 2005. Retrieved 9 July 2016.
- . Wikipedia. 2022-07-18 (英语).
- Sillero-Zubiri, Claudio; Hoffmann, Michael; Macdonald, David W.; IUCN/SSC Canid Specialist Group; IUCN--The World Conservation Union. . Gland, Switzerland. 2004 [2022-07-19]. ISBN 2-8317-0786-2. OCLC 57570754. (原始内容存档于2022-02-12).
- Macdonald, David W. . Unwin Hyman. 1989. ISBN 0-04-440419-0. OCLC 19776630.
- Nowak, Ronald M. Sixth. Baltimore: Johns Hopkins University Press. 1999: 636. ISBN 0-8018-5789-9. OCLC 39045218.
- Burnie, David; Wilson, Don E. 1st American. New York: DK. 2001. ISBN 0-7894-7764-5. OCLC 46422124.
- . BBC News. 2012-03-05 [2022-07-19]. (原始内容存档于2022-02-11) (英国英语).
- Klir, John J.; Heath, James E. . Physiological Zoology. 1992-09-01, 65 (5) [2022-07-20]. ISSN 0031-935X. doi:10.1086/physzool.65.5.30158555. (原始内容存档于2022-01-31).
- Larivière, Serge & Pasitschniak-Arts, Maria (1996). Vulpes vulpes (页面存档备份,存于) (PDF). American Society of Mammalogists.
- Nasimovich, A. A.; Bannikov, A. G.; Hoffmann, Robert S. . Leiden: Brill. 1989. ISBN 90-04-08873-3. OCLC 17953816.
- Albone, E. S. & Grönnerberg, T. O. "Lipids of the anal sac secretions of the red fox, Vulpes vulpes and of the lion, Panthera leo". Journal of Lipid Research. 18.4 (1977): 474–479.
- Fawcett, John K.; Fawcett, Jeanne M.; Soulsbury, Carl D. . Journal of Ethology. 2013-01-01, 31 (1). ISSN 1439-5444. doi:10.1007/s10164-012-0348-7 (英语).
- Macdonald, D. W. (1979). "Some observations and field experiments on the urine marking behaviour of the red fox, Vulpes vulpes L (页面存档备份,存于).". Zeitschrift für Tierpsychologie. 51.1: 1–22.
- Dahlstrøm, Preben; Walters, Martin. . Oxford: Oxford University Press. 2001. ISBN 0-19-850796-8. OCLC 45008608.
- Henry, J. David. . Behaviour. 1977-01-01, 61 (1-2) [2022-07-20]. ISSN 0005-7959. doi:10.1163/156853977X00496. (原始内容存档于2022-07-20) (英语).
- Andersen, , K.F.; Vulpius, T. . Chemical Senses. 1999-01-01, 24 (2). ISSN 0379-864X. doi:10.1093/chemse/24.2.179.
- Evans, Jonah; Kresky, Michael. . Berkeley: University of California Press. 2012. ISBN 978-0-520-95164-8. OCLC 802047175.
- Feldhamer, George A.; Thompson, Bruce Carlyle; Chapman, Joseph A. 2nd. Baltimore, Md.: Johns Hopkins University Press. 2003. ISBN 0-8018-7416-5. OCLC 51969059.
- Iossa, Graziella; Soulsbury, Carl D.; Baker, Philip J.; Harris, Stephen. . Journal of Mammalogy. 2008-12-16, 89 (6). ISSN 0022-2372. doi:10.1644/07-MAMM-A-405.1.
- Barrett, Priscilla. . Princeton, NJ: Princeton University Press. 2011. ISBN 978-0-691-15228-8. OCLC 724663784.
- Holland, Jennifer S. (July 2011). "40 winks?". National Geographic. 220 (1).
- , [2022-07-24], (原始内容存档于2020-08-31)
- . Psychology Today. [2021-02-05] (英国英语).
- . Bored Panda. [2021-02-05]. (原始内容存档于2022-11-26) (美国英语).
- Asa, C. S. & Mech, D. (1995). "A review of the sensory organs in wolves and their importance to life history," in Ecology and Conservation of Wolves in a Changing World eds. Carbyn, L. D.; Fritts, S. H. & Seip, D. R. (Edmonton: Canadian Circumpolar Institute): 287–291.
- Osterholm, H. (1964). "The significance of distance reception in the feeding behaviour of fox (Vulpes vulpes L.)". Acta Zoologica Fennica. 106 1–31.
- Wells, Michael C. . Behavioural Processes. 1978-07-01, 3 (2). ISSN 0376-6357. doi:10.1016/0376-6357(78)90041-4 (英语).
- Wells, Michael C.; Lehner, Philip N. . Animal Behaviour. 1978-02-01, 26. ISSN 0003-3472. doi:10.1016/0003-3472(78)90025-8 (英语).
- IUCN. . 2016-03-01 [2022-07-25]. doi:10.2305/iucn.uk.2021-1.rlts.t23062a193903628.en. (原始内容存档于2021-08-13) (英语).
- Mech, L. David; Boitani, Luigi. . Chicago: University of Chicago Press. 2003: 269. ISBN 0-226-51696-2. OCLC 51810720.
- Sargeant, Alan B.; Allen, Stephen H. . Journal of Mammalogy. 1989-08-18, 70 (3). ISSN 0022-2372. doi:10.2307/1381437.
- Scheinin, Shani; Yom-Tov, Yoram; Motro, Uzi; Geffen, Eli. . Animal Behaviour. 2006-03-01, 71 (3). ISSN 0003-3472. doi:10.1016/j.anbehav.2005.05.022 (英语).
- Nasimovich, A. A.; Bannikov, A. G.; Hoffmann, Robert S. . Leiden: Brill. 1989: 1278. ISBN 90-04-08873-3. OCLC 17953816.
- Nasimovich, A. A.; Bannikov, A. G.; Hoffmann, Robert S. . Leiden: Brill. 1989: 952. ISBN 90-04-08873-3. OCLC 17953816.
- Lewis, Deane. . The Owl Pages. [2022-07-26]. (原始内容存档于2022-10-25) (英语).
- Jaksić, F. M., & Marti, C. D. (1984). Comparative food habits of Bubo owls in Mediterranean-type ecosystems. Condor, 288-296.
- Watson, Jeff. 2nd. A&C Black. 2010: 92. ISBN 978-1-4081-1420-9.
- Lewis, C. F. (1957). Wedge-tailed eagle takes a fox. Victorian Naturalist, 74, 89-90.
- Utekhina, I., Potapov, E., & McGRADY, M. J. (2000). Diet of the Steller's Sea Eagle in the northern Sea of Okhotsk, pp. 71–92 in First Symposium on Steller's and White-tailed Sea Eagles in East Asia. Tokyo, Japan: Wild Bird Society of Japan.
- Larivière, S., & Pasitschniak-Arts, M. (1996). Vulpes vulpes. Mammalian species, (537), 1-11.
- Lalošević, Vesna; Lalošević, Dušan; Čapo, Ivan; Simin, Verica; Galfi, Annamaria; Traversa, Donato. . Parasite. 2013, 20 [2022-07-26]. ISSN 1776-1042. PMC 3718516 . PMID 23340229. doi:10.1051/parasite/2012003. (原始内容存档于2022-10-30) (英语).
- SMITH, H. J. . Journal of Wildlife Diseases. 1978-07-01, 14 (3). ISSN 0090-3558. doi:10.7589/0090-3558-14.3.366.
- Ancient Greek: Τευμησ(σ)ία ἀλώπηξ (Teumēs(s)íā alôpēx),gen.: Τευμησίας ἀλώπεκος, also known as ἀλώπηξ τῆς Τευμησσοῦ "fox of Teumessos"; Teumessos was an ancient city in Boeotia.
- Wallen, Martin. . London. 2006: 40. ISBN 978-1-86189-297-3. OCLC 67375486.
- Monaghan, Patricia. . New York: Facts On File. 2004: 199–200 [2022-07-26]. ISBN 0-8160-4524-0. OCLC 51886379. (原始内容存档于2022-05-24).
- Goff, Janet. (PDF). Japan Quarterly. 1997, 44 (2) [28 July 2012]. (原始内容 (PDF)存档于21 September 2013).
- Smyers, Karen Ann. . Honolulu: University of Hawaii Press. 1999. ISBN 0-8248-2058-4. OCLC 39523475.
- . Bible Gateway. [2022-07-26]. (原始内容存档于2022-11-05) (英语).
- . Bible Gateway. [2022-07-26]. (原始内容存档于2022-11-05) (英语).
- Bright, Michael. . London: Robson. 2006: 20–127. ISBN 1-86105-831-4. OCLC 59356395.
- Mitchell, Judy K. . Santa Barbara, Calif. 2004: 99–100 [2022-07-26]. ISBN 1-85109-538-1. OCLC 57190084. (原始内容存档于2020-08-24).
- Potts, Allen. . Washington: The Carnahan Press. 1912: 7, 38 [9 July 2016].
- . Trapping Today. [2022-07-28]. (原始内容存档于2022-11-05) (美国英语).
- . ghostarchive.org. [2022-07-28]. 原始内容存档于2022-01-09.
- Karim, Fariha. . [2022-07-28]. ISSN 0140-0460. (原始内容存档于2022-11-05) (英语).
- Trut, Lyudmila. . American Scientist. 1999, 87 (2) [2022-07-28]. ISSN 0003-0996. doi:10.1511/1999.2.160. (原始内容存档于2017-04-01) (英语).
- . web.archive.org. 2013-09-16 [2022-07-29]. 原始内容存档于2013-09-16.
- Troughton, Guy. . London: Whittet Books. 1986. ISBN 0-905483-47-2. OCLC 16645358.
- . www.wildsingapore.com. [2022-07-29]. (原始内容存档于2022-10-30).
- , [2022-07-29], (原始内容存档于2022-11-05) (中文(中国大陆))
- . BBC. [2022-07-29]. (原始内容存档于2022-11-05) (英国英语).
- . BBC News. [2022-07-29]. (原始内容存档于2022-11-05) (英国英语).
外部链接
| 维基共享资源中相关的多媒体资源:赤狐 |
| 維基物種上的相關:赤狐 |
- . ITIS. [18 March 2006].
- Red Fox (页面存档备份,存于), National Geographic
- Natural History of the Red Fox (页面存档备份,存于), Wildlife Online
- Sacramento Valley red fox info1 (页面存档备份,存于),
- A video of a family of red fox cubs, showing their behaviour around the den (页面存档备份,存于)
- A close up video of an adult male fox (页面存档备份,存于)
- Wild red fox hiding in woods and walking away (页面存档备份,存于)
- Red Fox (页面存档备份,存于), Fletcher Wildlife Garden
- 赤狐骨头的图片 (页面存档备份,存于)