耐梅蓋特母龍屬
耐梅蓋特母龍(屬名:Nemegtomaia,意為「耐梅蓋特的好母親」),是一屬偷蛋龍科的獸腳亞目恐龍,生存於白堊紀晚期的蒙古。正模標本發現於1996年,並成為命名新屬新種的依據,模式種巴氏耐梅蓋特母龍(N. barsboldi)於2004年發表。最初採用的屬名是Nemegtia,但已被一種介形亞綱所先使用,於是在2005年更名為Nemegtomaia。屬名中的Nemegt取自發現地的耐梅蓋特盆地,而maia有「好媽媽」的意思,象徵著偷蛋龍科會孵育牠們蛋的事實;種名紀念古生物學家瑞欽·巴思缽(Rinchen Barsbold)。2007年發現了另外兩具標本,其中一具是在巢穴上方與蛋一同被發現的,但本屬已在這項發現之前獲得這樣的屬名。
| 耐梅盖特母龙属 化石时期:白堊紀晚期 | |
|---|---|
 | |
| 重建骨架,位於河南地質博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 獸腳亞目 Theropoda |
| 科: | †偷蛋龍科 Oviraptoridae |
| 亚科: | †河源龙亚科 Heyunanniinae |
| 属: | †耐梅盖特母龙属 Nemegtomaia Lü et al., 2005 |
| 模式種 | |
| †巴氏耐梅蓋特母龍 Nemegtomaia barsboldi (Lü et al., 2004) | |
| 異名 | |
| |
耐梅蓋特母龍估計身長2公尺和體重40公斤。身為一種偷蛋龍類,牠可能長有羽毛。頭骨深、窄而短,頭頂上有一個拱形頭冠。沒有牙齒、短吻部有類似鸚鵡的嘴喙,並在齶上有一對牙齒狀突起。三只手指,其中第一指最大並長有強壯的爪子。耐梅蓋特母龍被分類為偷蛋龍科中的河源龍亞科,且是這個類群中已知唯一具有頭冠的物種。雖然耐梅蓋特母龍曾被拿來假設偷蛋龍科是一群不具飛行能力的鳥類,但普遍仍認為牠們是一群非鳥類恐龍。
連帶蛋巢的耐梅蓋特母龍標本坐在可能呈環狀排列的蛋上,並將雙臂環抱於其周圍。這些蛋沒有一個是完整的,但經測量在完好時寬約5至6公分及長約14至16公分。標本發現於沉積相地層,代表著牠們喜歡將巢穴建在接近河流的地區,那裡提供了柔軟、多沙的基質和豐沛的食物來源。牠們可能透過將尾羽和翼羽覆蓋在蛋上來提供保護。標本的骨骼已被損壞,呈現出被皮蠹科所啃食的跡象。偷蛋龍科的食性目前尚未明瞭,但牠們的頭骨與其他也是、或至少被認為是植食性動物的最為相似。耐梅蓋特母龍的化石來自耐梅蓋特盆地的西戈約特組,當地環境被認為同時具有乾旱兼潮濕的特性。
發現歷史
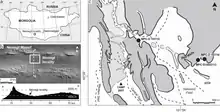
1996年,日本古生物學家小林快次(「蒙古高原國際恐龍計畫」團隊的一員)在蒙古西南部戈壁沙漠的耐梅蓋特組發現了一具不完整的偷蛋龍科骨骼。標本(編號MPC-D 100/2112,屬於蒙古古生物學中心,之前是PC和GIN 100/2112)由一個幾乎完整的頭骨和部分顱後骨骼(包含頸椎、胸椎、薦骨、尾椎、左肩胛骨、成對肱骨的下端、右橈骨、成對的髂骨、成對恥骨的上端、成對的坐骨、以及一個股骨的上端)所組成。2002年中國古生物學家呂君昌和同僚將標本敘述為機靈龍的新未命名種(Ingenia sp.),並用來強調偷蛋龍類與鳥類間的相似性。[1][2][3]

B-D.巢穴標本覆蓋於蛋上的骨骼
E-G.下層的蛋
2004年,呂君昌和同事確定骨骼屬於一個新的獨立物種,並將其作為巴氏耐梅蓋特龍(Nemegtia barsboldi)的正模標本。屬名意指「來自耐梅蓋特盆地」;種名紀念蒙古古生物學家瑞欽·巴思缽(Rinchen Barsbold),他也是發現標本團隊的領導人。[1]2005年,敘述者在經過生物學家的提醒後發現,Nemegtia的屬名已被一種來自同一地層、於1978年命名的介形綱所優先使用,於是他們提出了新的屬名Nemegtomaia(加上希臘文的maia使全名意為「耐梅蓋特的好母親」),象徵著當時的新發現:偷蛋龍科會孵蛋而不是偷蛋,雖然這時候還沒發現與耐梅蓋特母龍有關的巢穴或蛋證據。[4][5]最早發現的偷蛋龍科,其巢穴最初被認為屬於原角龍,因此於1924年命名成偷蛋龍,意為「偷蛋的盜賊」。直到1990年代,有更多連帶巢穴和蛋的偷蛋龍科標本被發現,根據保存下來的胚胎,證明這是偷蛋龍科自己的蛋。[6][7][8]相似的更名情況也在機靈龍(Ingenia)出現,因為屬名被一種線蟲動物門先使用,於2013年更名為雌駝龍(Ajancingenia)。[9]

2007年,由戈壁恐龍考察團隊發現了兩具新的耐梅蓋特母龍標本,並於2013年由義大利古生物學家費德里科·芬蒂(Federico Fanti)和同僚所敘述。第一件標本MPC-D 107/15由芬蒂在西戈約特組發現,他將其暱稱為「Mary」,包含一個蛋巢以及上方一具推測是親屬的骨骼。作為偷蛋龍科中第四個發現於巢穴之上的物種(排在偷蛋龍、葬火龍、以及曲劍龍之後),耐梅蓋特母龍在這個發現之前即獲得了與這個形象相符的名稱。這個標本是在「艱困的情況下」從懸崖壁上挖掘出來,還遇到了大雨和砂岩坍塌等困難。[2][5]
巢穴的骨骼保存了部分頭骨、成對肩胛骨、左臂和手、右肱骨、恥骨、坐骨、股骨、脛骨、腓骨、兩隻腳的下部。標本在距離正模標本不到500公尺處發現,且體型相同;根據相似的生理解剖特徵、地理位置接近而被歸入耐梅蓋特母龍。採集時將骨骼和巢穴包覆在同一個岩塊之中,藉此保存好彼此在空間上的關聯性。第二件標本MPC-D 107/16由美國古生物學家尼克·朗里奇(Nicholas R. Longrich)在耐梅蓋特組發現,包含手部、部分左尺骨和橈骨、肋骨、部分骨盆、以及成對的股骨。這個標本比其他的小了約35%,並根據與MPC-D 107/15標本相似的手部特徵而歸入耐梅蓋特母龍。這些手部可能屬於不同的個體,因為它們並未和其他骨骼關節連接在一起(同採集地還發現過其他偷蛋龍科的多具骨骼),但至今仍無法確認。[2]
敘述

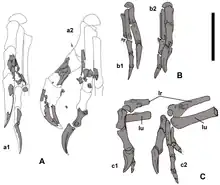
耐梅蓋特母龍經測量身長約2公尺,體重約40公斤,這是根據已知更完整的近緣物種所推測出來的體型。作為一種偷蛋龍類,牠可能擁有羽毛。頸椎神經棘短,而神經弓外觀呈X字形。中間三節頸椎最大。肩胛骨全長185毫米。肱骨長152毫米,在與現代鳥類相同位置有一個凹孔,但這在偷蛋龍類中並不尋常。橈骨直,剖面為卵圓形,並可能長144毫米。第一個手指相當大並長有一根強壯的爪子,且比另外兩個手指還要大。第二個手指比第一個略長,而第三個手指最短小。骨盆髂骨的上緣筆直,雖然兩個髂骨彼此靠近卻未融合在一起。恥骨軸向後扭轉。測量股骨長286毫米、脛骨長317毫米。[1][2]
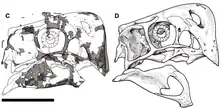
耐梅蓋特母龍的頭骨深、狹窄且短(與身體其餘部分比較),長度達到179毫米。有一個發育良好的頭冠,由吻部的鼻骨和前上頜骨所構成。正模標本頭冠的前緣與頭骨上緣幾乎垂直,呈90度角。與其他偷蛋龍科比較,前上頜骨的鼻突從上面視角幾乎看不到(這在頭冠最高點與鼻骨接觸)。頭冠向後及向下延伸,在最高點構成一個拱形輪廓。眼眶直徑為52毫米;因為頭骨短而使眼睛看起來很大。眼睛前面的腔室由兩個孔洞組成:後方是一個大的眶前孔、前方是一個小的上頜孔。耐梅蓋特母龍和其他偷蛋龍科的差異在於,頭骨中線的額骨長度為頂骨前後長度的25%。鼻孔相對小,位於頭骨上位。[1][2]

耐梅蓋特母龍的下頜無齒,而且就像其他偷蛋龍科,吻部短,帶有一個深、結實、某種程度上形似鸚鵡的嘴喙。有一個由前上頜骨、犁骨和上頜骨所構成的硬齶,如同其他偷蛋龍科。齶部朝下強烈彎曲,中間有一道裂痕。如同其他偷蛋龍科,齶上有一對朝下的牙齒狀突起(被稱作「偽齒」)。耐梅蓋特母龍吻部前面的前上頜骨之間的縫線(關節)側邊有小孔洞,可能代表著角蛋白質鞘的存在。下頜短且深,下面凸起,長153毫米。齒骨最高點達到50毫米。下頜骨縫(兩塊下頜骨連接處)短、深、而且高度氣腔化(具有空腔)。下頜孔大並位於下頜前部。[1][2][3][10]如同其他偷蛋龍科,下頜前部向下彎曲。[11]
雖然耐梅蓋特母龍並沒有單一項獨特的鑑定特徵(自衍徵)與其他偷蛋龍科區別開來,但頭冠、加大的第一指、薦骨數量多(8個)的特徵組合是本屬的獨特之處。[12]
分類
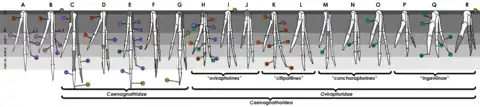
在2004年的系統發生學分析中,呂君昌和同事將耐梅蓋特母龍分類成一種進階型的偷蛋龍類,並發現牠和葬火龍親緣關係最近。2010年朗里奇和同事確認耐梅蓋特母龍屬於偷蛋龍科中,機靈龍亞科(Ingeniinae)的一部分,使牠成為後者中唯一具有顯著頭冠的物種。另一個偷蛋龍亞科的物種全部都有頭冠。機靈龍亞科的物種以較小的體型、短而健壯的前肢、輕微彎曲的指爪、綜薦骨所含的脊椎數目、以及腳部和骨盆的決定性特徵為鑑定特色。朗里奇和同事認為耐梅蓋特母龍的頭冠可能代表著這項特徵在偷蛋龍科中演化出來或消失了好幾次,或是牠根本不屬於機靈龍亞科。[2][13]2010年葛瑞格利·保羅(Gregory S. Paul)認為沒有頭冠的偷蛋龍科是其他有頭冠物種的幼年或雌性個體,而科中的物種數是被過度高估。他將耐梅蓋特母龍列為巴氏葬火龍("Citipati" (=Nemegtomaia) barsboldi),認為本種和葬火龍屬非常相似。但後來2016年他將其改列為巴氏竊螺龍("Conchoraptor" (=Nemegtomaia) barsboldi)。[10][14]
2012年芬蒂和同事也發現耐梅蓋特母龍屬於機靈龍亞科的進階型物種,並根據兩件新標本的手部比例(相對短和健壯的第一指),認為與河源龍近緣。他們表示雖然頭冠的存在通常與偷蛋龍亞科而非機靈龍亞科有關連,這個特徵可能與體型和成熟度有關。並指出機靈龍亞科的竊螺龍的鼻骨和額骨氣腔化,當個體成熟後可能有潛力長出頭冠,儘管該屬所有已知骨骼體型都一樣小(其中一個已完全成長)。[2]機靈龍亞科因為學名重複,後來由河源龍亞科取而代之。[15]以下演化樹取自芬蒂等人(2012),顯示耐梅蓋特母龍在偷蛋龍科中的位置:[2]

| 偷蛋龍科 Oviraptoridae |
| |||||||||||||||||||||||||||||||||||||||||||||
演化
偷蛋龍下目普遍被認為是一群非鳥類的獸腳亞目恐龍,而牠們與鳥類的相似性經常被拿來比較。近頜龍超科的化石在歷史上常與鳥類的混淆,因此某些研究人員開始認為偷蛋龍類其實與鳥類的關係比與非鳥類恐龍的關係更接近。2002年呂君昌和同事使用當時尚未命名的耐梅蓋特母龍作為鳥類和偷蛋龍類相似性比較,並發現後者實際上比一些類似鳥類的恐龍(如馳龍科)還要接近鳥類。他們更提出偷蛋龍類是一群不具飛行能力的鳥類而不是非鳥類恐龍,並強調恐龍和鳥類之間的分界越來越難以描繪。[3]其他學者則認為馳龍科和傷齒龍科最接近鳥類,牠們一同構成近鳥類演化支;而偷蛋龍類、鐮刀龍類、阿瓦拉慈龍類被排除於其外。包含偷蛋龍類和近鳥類在內的更大範圍演化支稱為廓羽盜龍類,根據有羽梗的羽毛來定義。[16]
已知的偷蛋龍類都來自亞洲(可能是起源地)和北美洲,並主要來自白堊紀晚期坎潘階至馬斯垂克階的地層。其中包含小型至大型物種,特徵為短的頭骨和喙、加長的手指和短的尾部。基群(或「原始」)物種具有牙齒,這在進階物種(近頜龍超科,包含偷蛋龍科)就消失了。牠們至少是部分的植食性動物,並用和鳥類一樣的姿勢來孵蛋。儘管牠們全部都被認為帶有羽毛,很明顯的無法飛行。頭冠似乎在類群中的不同族系間趨同演化。[16]耐梅蓋特母龍所屬的偷蛋龍科由普遍較小型的物種組成,且所知僅限於上白堊紀的亞洲,大部分物種發現於蒙古和中國的戈壁沙漠。[13]包含耐梅蓋特母龍在內,在戈壁沙漠的一個相對較小的地理區中至少發現了九屬偷蛋龍科。[12][17]
古生物學
繁殖

耐梅蓋特母龍的MPC-D 107/15標本與蛋巢一同被發現;牠把腳放在可能呈環狀排列的蛋列中央,雙臂交叉於身體兩側的蛋上,與其他孵巢偷蛋龍科化石呈現同樣姿勢。採集到的巢穴部分寬90公分、長100公分;骨骼佔整塊的上部25公分,其餘20公分由碎裂的蛋及蛋殼佔據。巢中沒有發現植物材料的證據,但有一些未定的骨骼碎片。巢沒留下任何完整的蛋或胚胎,使得巢中蛋的尺寸、形狀、數量和排列方式難以鑑定。可能是原本身體下方有兩層蛋,而巢中央沒有蛋。大部分的蛋(已鑑定出各別的7顆)和蛋碎片都在巢穴下層或標本頭骨、頸部、四肢底下發現,而骨骼是直接坐落在蛋上或其表面5毫米處。骨骼直接位在巢頂代表巢穴未被沙子覆蓋。雖然蛋的位置無法看出巢中的排列方式,但根據其他偷蛋龍科巢穴顯示出蛋是成對排列,並至少排列出三個同心圓。MPC-D 107/15的蛋可能在埋葬過程中受到強風、沙塵暴或掠食者等外來因素而移位。這也支持了蛋上方未有沙子掩埋的理論,因為完全掩埋的蛋不太可能受到外力而位移。[2]

偷蛋龍科的蛋平均長度17公分,而MPC-D 107/15標本中最完整的蛋被認為原本寬5至6公分及長14至16公分。這些蛋與過去在蒙古發現的長蛋龍科蛋化石幾乎一致。蛋殼相對薄,界於1至1.2毫米,表面覆蓋著隆起約0.3毫米的嵴節。蛋殼的顯微鏡學無法被妥善的研究,因為受到方解石的大幅重組和再結晶 (地質學)。[2] 孵巢標本在沉積區發現,代表偷蛋龍科偏好在河流附近築巢,此地在乾燥環境(濕氣少)中提供柔軟的沙質基質和食物。許多偷蛋龍科被發現維持在孵蛋姿勢,代表牠們可能需要較長時間來孵蛋,類似鴕鳥、鴯鶓和黑胸禿鷹等現代鳥類,牠們孵卵超過40天且提供有限的營養。在沙漠環境中築巢可能會因為高熱,而對白天大多時間停留在巢中的成年孵育者、蛋和幼雛造成傷害。因此築巢地點的選擇可能是在極端高溫下孵育的一種對抗機制。也有認為偷蛋龍科的尾羽是提供蛋巢遮蔽和保護的一種適應性構造。河源龍亞科的第二指與健壯的第一指相比尺寸明顯縮小也是功能改變的一種解釋,可能與第二指上附著的長翼羽有關。這些翼羽可能在孵蛋時提供保護。一旦第二指開始作為羽毛支撐,抓握能力便隨之降低,並由第一指取代而變得更加堅實。第三指也可能因為擋在翼羽後面,無法有效抓握而縮小。[2]
2018年台灣古生物學家楊子睿和同事檢驗手盜龍類(包含偷蛋龍類)的蛋殼皮層。在現代鳥類中,這種皮層(主要由脂類和羥磷灰石構成)能夠保護蛋免受脫水及微生物侵害。研究人員認為皮質蛋是一項適應性特徵,使耐梅蓋特母龍和其他偷蛋龍類在變動的環境中提高孵育成功率。[18]

許多研究指出許多隻雌性會將蛋一起產在同一個巢穴中,並排列好以便讓一隻個體(可能是雄性)來保護。[2]2010年美國古生物學家大衛·瓦立丘(David J. Varricchio)和同事發現偷蛋龍科和傷齒龍科相對大的窩卵數與一些現存主龍類(鳥類和鱷魚,與恐龍最近緣的現存動物)實施的多配偶制和普遍由雄性來提供親代撫育非常類似(也可在古顎類如鴕鳥、鴯鶓中觀察到)。這樣的生殖系統出現早於鳥類的起源,而成為現代鳥類的祖先,雙親撫育則是後來才發展出來的。[19]
許多偷蛋龍類尾部末端都有尾綜骨,代表扇狀羽的存在;美國古生物學家W. Scott Persons和同事於2013年提出這項構造可能用於動物交流,例如求偶展示。[20]
食性

自從最初偷蛋龍被錯誤描述為偷蛋的掠食者以來,關於牠們的食性已經提出許多種不同的看法。根據在尾羽龍中發現的胃石、還有門齒龍的牙齒磨損面,推測所有的偷蛋龍類都是植食性。2010年朗里奇和同事發現偷蛋龍科下頜具有一些在植食性四足類(尤其是二齒獸下目,一群已滅絕的合弓綱似哺乳爬行動物)所能觀察到的特徵。偷蛋龍科和二齒獸類的共有特徵包含:短而深、無齒的下頜;加長的齒骨聯合;加長的下頜孔;齶部有道朝下的骨板。現代動物中下頜與偷蛋龍科類似的有鸚鵡和陸龜;後者的前上頜骨也有齒狀突。朗里奇和同事得出結論:因為偷蛋龍科和植食性動物的相似性,牠們大部分食物組成可能為植物。在地層中發現偷蛋龍科的頻率也很高,一如其他植食性恐龍的分布模式;這些動物會比肉食性的還要繁盛,因為牠們位在食物鏈中較低階的營養級,可獲得較充足的食物能量。偷蛋龍科的下頜可能適用於處理食物,例如生長在其生活環境的耐旱植物(適應乾燥缺水的環境),但這點無法確認,因為缺乏對當時植被組成的了解。[13]2013年呂君昌和同事的研究發現偷蛋龍科可能在成長過程中維持後肢的比例,也如同植食性動物的成長模式。[21]2017年,加拿大古生物學家葛瑞戈里·方斯頓(Gregory F. Funston)和同事認為,偷蛋龍科鸚鵡似的嘴喙,可能代表著一種以堅果和種子為食的食果習性。[12]
1977年巴思缽提出偷蛋龍科是種食螺性動物,但朗里奇和同事反駁這項論點,他們透過模擬食殼性動物壓碎硬殼的方式,發現這類動物通常需要具有較寬的牙齒面積來壓碎。然而偷蛋龍科齒骨形狀表明牠們具有鋒利的喙,適合切割堅硬的食物,而不是用來打碎堅硬材質的貝殼或蛋殼。齒骨前部的融合板可能有一定程度的壓碎能力,但面積相對較小,可能不是作為頜部的主要功能。大部分發現偷蛋龍科的沉積層也被解釋為乾燥或半乾燥環境,再次反駁專食貝類和蛋的論點,因為在這樣的氣候條件下無法提供足夠的食物支持量。[13]
朗里奇和同事指出,河源龍亞科健壯的前肢和加大的第一指與一些現代食蟻性動物(如食蟻獸、穿山甲)相似,但河源龍亞科的下頜形態不足以支持牠們是食蟲動物。研究人員發現河源龍亞科的前肢功能不明瞭,但推測可能用來刮取、撕扯或挖掘,此外並不包含捕捉獵物。[13]
2004年呂君昌和同事提出,根據耐梅蓋特母龍方骨與方軛骨間的關節,代表這些彼此相連的骨頭可以作頭骨運動,影響到頜部功能。2015年比利時古生物學家克里斯托夫·亨德里克斯(Christophe Hendrickx)和同事發現耐梅蓋特母龍的頭骨不太可能作出如鳥類般的運動,因為方骨是無法移動的。[22]
古生態學

耐梅蓋特母龍來自耐梅蓋特組及西戈約特組,地質年代可追溯至白堊紀晚期的上坎潘階至下馬斯垂克階,約7000萬年前。雖然對於本屬的所知僅來自耐梅蓋特化石點,一些來自其他化石點的未確認偷蛋龍科遺骸也可能屬於耐梅蓋特母龍。耐梅蓋特山塊有許多山谷和峽谷,有些深達40公尺,其中有最良好的地層暴露出來。耐梅蓋特組的岩相呈現出一個潮濕的河流作用環境,而西戈約特組則表現出一個乾旱或半乾旱環境以及風蝕作用的岩床。這兩個地層與其中多樣的化石,從時間上來看被認為代表著由具有差異的氣候環境所組成的連續時段,但2009年加拿大古生物學家大衛·埃伯斯(David A. Eberth)和同事發現兩者交界處有部分重疊,約有25公尺厚的交插(interfinger)狀態,表示當地在沉積過程時,河流及風蝕作用環境是同時存在的。[2][12][23][24]

耐梅蓋特組的環境被拿來與現今波札那的歐卡萬哥三角洲相提並論。[25]耐梅蓋特河中及周圍環境為多樣化的生物提供了棲地。水生動物包含軟體動物、魚類及烏龜、鱷形超目的沙漠鱷屬。已發現的哺乳類化石有多瘤齒獸目;鳥類則有格日勒鳥、朱迪鳥、特維鳥。植食性恐龍包含:甲龍科的多智龍;厚頭龍科的傾頭龍;鴨嘴龍科的櫛龍和巴思缽氏龍;蜥腳類的納摩蓋吐龍和後凹尾龍。其他獸腳類恐龍包含:暴龍超科的特暴龍、分支龍、小掠龍;傷齒龍科的無聊龍、鴕鳥龍、蜥鳥龍;鐮刀龍科的鐮刀龍;似鳥龍類的恐手龍、似鵝龍、似雞龍。[26][27][28]
其他來自耐梅蓋特組的偷蛋龍類包含:基礎的擬鳥龍;偷蛋龍科的瑞欽龍、天青石龍、竊螺龍和雌駝龍;近頜龍科的單足龍。雖然當地的偷蛋龍類物種數很多(耐梅蓋特組在任何方面都具有最高的偷蛋龍類多樣性),卻沒有任何一種是彼此近緣。耐梅蓋特組的獨特之處在於偷蛋龍科和近頜龍科同時存在;1993年加拿大古生物學家菲力·柯里(Phillip J. Currie)和同事認為這種多樣性源自於兩種偏好不同環境的族群;2016年日本古生物學家對比地孝亘和同事認為,根據兩者發現於不同的地層,偷蛋龍科可能偏好較乾燥的環境,而近頜龍科偏好河相環境。[2][29]方斯頓和同事發現偷蛋龍科同時出產於乾燥和中濕棲地(但前者較為繁盛),而其他偷蛋龍類避免居住在乾燥環境,且類群共存的現象可用食性上的棲位分化來解釋。耐梅蓋特組產生如綠洲般的拉力或誘因,吸引偷蛋龍科來此居住。[12]
埋藏學

孵巢標本MPC-D 107/15提供了許多關於西戈約特組埋藏過程(腐爛和化石化時的變化)的資訊。標本保存於被沙塵暴或沙丘位移的沉積相。似乎未有死後被移運的跡象,但屍體輕微向右偏轉,表示沉積物來自左邊。頸部向左彎曲,左臂向後折疊,雙腿屈膝蹲下。埋藏過程中脊柱、頸部及骨盆受損,而骨骼的大部分損壞被認為是由無脊椎動物活動所造成。[2]
標本的骨頭碎屑、孔洞、改造沉積物(可能因為結蛹造成)指出屍體曾被皮蠹科或其他食腐昆蟲啃食。骨骼關節處有許多啃食的痕跡,且幾乎所有骨骼連接面都被損壞。頭部和頸部下方的巢穴也有類似的通道痕跡,在這些痕跡處也沒找到任何蛋的殘留物。現代的皮蠹主要以肌肉組織為食,但會避開潮濕物質,受到快速掩埋時其活動也會被阻礙。所以可以推測MPC-D 107/15標本起初僅被部分掩埋,上部暴露在外而引來一群皮蠹。某些骨骼受損(尤其是脊柱部分)也可能是由小型哺乳動物的食腐所造成。[2]
參考來源
- Lü, J.; Tomida, Y.; Azuma, Y.; Dong, Z.; Lee, Y.-N. (PDF). Bulletin of the National Science Museum, Tokyo, Series C. 2004, 30: 95–130 [2020-12-13]. (原始内容 (PDF)存档于2021-08-31).
- Fanti, F.; Currie, P. J.; Badamgarav, D.; Lalueza-Fox, C. . PLOS ONE. 2012, 7 (2): e31330. Bibcode:2012PLoSO...731330F. PMC 3275628
 . PMID 22347465. doi:10.1371/journal.pone.0031330.
. PMID 22347465. doi:10.1371/journal.pone.0031330. - Lü, J., Dong, Z., Azuma, Y., Barsbold, R. & Tomida, Y. (2002). "Oviraptorosaurs compared to birds." In Zhou, Z. & Zhang, F. (eds). Proceedings of the 5th Symposium of the Society of Avian Paleontology and Evolution. Beijing: Science Press. pp. 175–189.
- Lü, J.; Tomida, Y.; Azuma, Y.; Dong, Z.; Lee, Y.-N.; et al. . Bulletin of the National Science Museum, Tokyo, Series C. 2005, 31: 51.
- Arbour, V.M. . pseudoplocephalus.blogspot.com. 2012 [2017-01-10]. (原始内容存档于2020-08-04).
- Norell, M.A.; Clark, J. M.; Chiappe, L.M.; Dashzeveg, D. . Nature. 1995, 378 (6559): 774–776. Bibcode:1995Natur.378..774N. doi:10.1038/378774a0.
- Dong, Z.-M.; Currie, P.J. . Canadian Journal of Earth Sciences. 1996, 33 (4): 631–636. Bibcode:1996CaJES..33..631D. doi:10.1139/e96-046.
- Clark, J. M.; Norell, M.; Chiappe, L. M. . American Museum Novitates. 1999, (3265): 1–35. hdl:2246/3102.
- Easter, J. . Zootaxa. 2013, 3737 (2): 184–190. PMID 25112747. doi:10.11646/zootaxa.3737.2.6.
- Paul, G. S. . Princeton University Press. 2010: 152, 154. ISBN 978-0-691-13720-9.
- Lü, J.; Yi, L.; Zhong, H.; Wei, X.; Dodson, P. . PLoS ONE. 2013, 8 (11): e80557. Bibcode:2013PLoSO...880557L. PMC 3842309 . PMID 24312233. doi:10.1371/journal.pone.0080557.
- Funston, G.F.; Mendonca, S.E.; Currie, P.J.; Barsbold, R. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2017, 494: 101–120. Bibcode:2018PPP...494..101F. doi:10.1016/j.palaeo.2017.10.023.
- Longrich, N.R.; Currie, P.J.; Zhi-Ming, D. . Palaeontology. 2010, 53 (5): 945–960. doi:10.1111/j.1475-4983.2010.00968.x.
- Paul, G. S. . Princeton University Press. 2016: 181. ISBN 978-0-691-16766-4.
- Yun, C.-G. . Zootaxa. 2019, 4671 (2): 295–296. PMID 31716062. doi:10.11646/zootaxa.4671.2.11.
- Hendrickx, C.; Hartman, S.A.; Mateus, O. . PalArch's Journal of Vertebrate Palaeontology. 2015, 12 (1): 1–73 [2020-12-13]. (原始内容存档于2018-06-22).
- Wang, S.; Sun, C.; Sullivan, C.; Xu, X. . Zootaxa. 2013, 3640 (2): 242–57. PMID 26000415. doi:10.11646/zootaxa.3640.2.7.
- Yang, T.-.; Chen, Y.-H.; Wiemann, J.; Spiering, B.; Sander, P. M. . PeerJ. 2018, 6: e5144. PMC 6037156 . PMID 30002976. doi:10.7717/peerj.5144.
- Varricchio, D.J.; Moore, J.R.; Erickson, G.M.; Norell, M.A.; Jackson, F.D.; Borkowski, J.J. . Science. 2008, 322 (5909): 1826–1828. Bibcode:2008Sci...322.1826V. PMID 19095938. doi:10.1126/science.1163245.
- Persons, S. . Acta Palaeontologica Polonica. 2013. doi:10.4202/app.2012.0093.
- Lü, J.; Currie, P.J.; Xu, L.; Zhang, X.; Pu, H.; Jia, S. . Naturwissenschaften. 2013, 100 (2): 165–175. Bibcode:2013NW....100..165L. PMID 23314810. doi:10.1007/s00114-012-1007-0.
- Hendrickx, C.; Araújo, R.; Mateus, O. . PeerJ. 2015, 3: e1245. PMC 4579021 . PMID 26401455. doi:10.7717/peerj.1245.
- Eberth, D. A.; Badamgarav, D.; Currie, P. J. . 고생물학회지. 2009, 25 (1) [2020-12-19]. ISSN 1225-0929. (原始内容存档于2018-06-12).
- Eberth, D.A. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2017, 494: 29–50. Bibcode:2018PPP...494...29E. doi:10.1016/j.palaeo.2017.11.018.
- Holtz, T.R. . Nature. 2014, 515 (7526): 203–205. Bibcode:2014Natur.515..203H. PMID 25337885. doi:10.1038/nature13930.
- Lucas, S.G.; Estep, J.W. . New Mexico Museum of Natural History and Science Bulletin. 1998, 14: 1–20 [2020-12-19]. (原始内容存档于2018-11-19).
- Watabe, M.; Tsogtbaatar, K.; Suzuki, S.; Saneyoshi, M. . Hayashibara Museum of Natural Sciences Research Bulletin. 2010, 3: 11–18.
- Jerzykiewicz, T.; Russell, D.A. . Cretaceous Research. 1991, 12 (4): 345–377. doi:10.1016/0195-6671(91)90015-5.
- Tsuihiji, T.; Watabe, M.; Tsogtbaatar, K.; Barsbold, R. . Cretaceous Research. 2016, 63: 148–153. doi:10.1016/j.cretres.2016.03.007.