螯肢亞門
螯肢亞門(學名:)是一類主要的節肢動物,其下包括鱟、蠍子、蜘蛛及蟎。它們可能是於寒武紀演化自海生動物。現存的水生物種包括劍尾目的4個物種及約1300種的海蜘蛛。陸地上已確認的螯肢亞門有超過7.7萬種,而未經確認的也有50萬種。
| 螯肢亚门 化石时期:寒武纪至現今 | |
|---|---|
 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 节肢动物门 Arthropoda |
| 演化支: | 蛛形类 Arachnomorpha |
| 亚门: | 螯肢亚门 Chelicerata Heymons, 1901 |
| 演化支 | |
螯肢亞門像其他節肢動物般身體有節,並由甲殼素及蛋白質組成的表皮覆蓋。它們的身體有兩個體區,即頭胸部及腹部,而蟎在這兩部份的分隔則不可見。它們口器前有一螯肢。大部份亞類都只會以螯肢來吃食,但蜘蛛螯肢的螯爪則形成了獠牙狀,大部份物種更可以從中注入毒素至獵物中。它們的循環系統是開放的,由管狀的心臟將血液經血腔輸送至身體其他部份。水生螯肢亞門是有鰓的,而其他的則有書肺及氣管。螯肢亞門中央神經系統的神經節融合成一團,融合的程度各有不同,如最為古老及原始的中突蛛亞目神經節的融合程度則很有限。大部份螯肢亞門都有剛毛來感應振動、氣流及化學轉變。最活躍的獵食性蜘蛛有非常銳利的視覺。
螯肢亞門最初只是掠食者,但後來分化出多種覓食策略,包括掠食、寄生、植食性、食腐性及腐生性。雖然盲蛛目可以消化固體食物,但其他螯肢亞門的食道太窄,一般只會用螯肢及消化酶將食物液化。陸生的螯肢亞門為了保存水份只會排泄固體。陸生的螯肢亞門一般都是體內受精,而水生的鱟則是體外受精的。掠食性的物種會採取一種很小心的求偶方法,以避免雄性在交配前被雌性吃掉。大部份物種在孵化後都會像成體,但所有蠍子及一些蟎都會將卵保存在其體內,直至幼體出生。大部份螯肢亞門的幼體都是自行覓食的,但雌性的蠍子及一些蜘蛛是會保護及餵飼幼體的。
螯肢亞門的起源一直都存在爭論。雖然對於亞類之間的關係有一定共識,但海蜘蛛的分類及蠍子的定位仍有爭議。[1]
雖然一些蜘蛛及蠍子的毒素非常危險,醫護界現正研究使用這些毒素來治療癌症至勃起功能障礙等病症。鱟的血液也有用來測試細菌的存在。基因工程師改造羊奶及葉子製造出蜘蛛絲。蟎可以令人過敏,並會傳播疾病,故是一種害蟲。
特徵
節及表皮

螯肢亞門的身體及肢體有節,並由甲殼素及蛋白質組成的表皮覆蓋。其頭部由幾節組成,這些節於胚胎發育時就已經融合在一起。體腔非常細小,血腔連接至管狀的心臟。[2]它們的身體分為兩個體區,最前端的頭胸部與及後方的腹部。[2]但是蜱蟎亞綱的頭胸部及腹部並沒有明顯的分野。[2]
螯肢亞門的頭胸部於胚胎時期透過融合帶有眼睛的口前葉和第二至第七節生成,第一節於胚胎發育期間會消失。第二節有一對螯肢,第三節有一對鬚肢,很多亞類都是以此來感官的,其餘的四節都有腳。較原始形態的口前葉兩側有一對複眼,中央有四隻單眼。[2]口器是在第二及第三節之間。[3]
腹部包含12或更少的節數,頭七節為前腹部,後五節為後腹部,以尾節或尖刺終結。現今螯肢亞門已經失去了前腹部附肢,或是已經大幅演化[2],如蜘蛛的噴絲板[2]或鱟的鰓。[2]
螯肢及鬚肢
螯肢亞門的螯肢及鬚肢在形態及功能上有很大分野,唯一相同的就是其位置:螯肢出於第竹二節,位於口器前;而鬚肢來自第三節,位於口器之後。[2]
螯肢一般分三節,第三節上有螯爪。[2][4]不過,蜘蛛的螯肢只有兩節,於第二節有螯爪,不使用時螯爪可以收起。[2]螯肢的大小也各有不同:板足鲎的螯爪很大[4],而蠍子的則很細小。[2]
大部份螯肢亞門的鬚肢相對較為細小及用作感官。[2]不過雄性蜘蛛的鬚肢端呈球狀,可以作為針筒將精子注入雌性的生殖口[2],而蠍子的則有大鉗用來捕捉獵物。[2]
體腔及循環系統
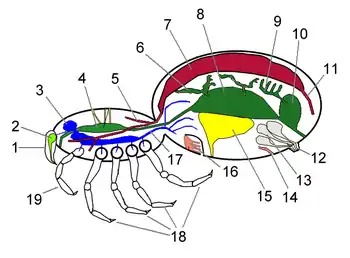 | |||||||||||||||||||||||||||||||||||||||||
蜘蛛的主要器官
| |||||||||||||||||||||||||||||||||||||||||
生理系統
| |||||||||||||||||||||||||||||||||||||||||
螯肢亞門的體腔非常細小,在生殖系統及排泄系統之外只餘少許空間。主體腔是將血液帶到全身的血腔,管狀的心臟會將後部的血液向前供血。雖然動脈會將血液帶到特定的身體部位,但末端是開放的,並沒有接合至靜脈,是為開放式的循環系統。[2]
呼吸系統
螯肢亞門的呼吸系統會就亞類所生存的環境而有所不同。現存陸生的螯肢亞門一般都有書肺及氣管,書肺會經血液供應氧及移除廢氣,而氣管可以不用經血液而發揮同樣功能。[2]鱟是水生的,故在類同位置有書鰓。已滅絕的板足鲎目估計是有鰓的,但其化石證明卻很混淆。但是奧陶紀晚期Onychopterella的化石似乎有三對書鰓,外觀非常像蠍子書肺。[5]
覓食及消化
大部份螯肢亞門的食道太窄,不足以容納固體食物。[2]所有蠍子及差不多所有蜘蛛都是掠食者,並會預先在口前腔用螯肢處理獵物。[2]不過有一種蜘蛛是植食性的[6],並有很多是會以花蜜及花粉來補充其營養。[7]大部份蜱蟎亞綱都是吸血的寄生蟲,但也有掠食性、植食性及食腐性的亞類。所有蜱蟎亞綱都有可收縮的螯肢、鬚肢及部份外骨骼,組成口前腔來預先處理食物。[2]
盲蛛目是少數現存螯肢亞門可以吃固體食物,當中也有掠食性、植食性及食腐性的。[2]鱟有獨特的消化系統可以處理固體食物。它們腳端的爪會將細小的無脊椎動物抓入最後肢至口器間的食物溝,腳基位置有鋸齒狀的顎基可以磨碎食物,並將之放入口內。[2]這種方式相信是最古老的節肢動物的覓食方法。[8]
排泄
鱟會將氮廢物轉為氨,再經鰓排出。其他的廢物會成為糞便由肛門排出,另外亦有腎管遣濾出尿液。[2]氨是有毒的,故它們需要大量水份來稀釋。[2]大部份陸生的螯肢亞門都不能消耗如此大量的水份,故會將氮轉為其他化合物,並以固體形式排出。排泄可以由腎管及馬氏管進行。這些管都是從血液中過濾廢物,並以固體形態排到後腸。[2]
神經系統
| 頭胸神經節融入至腦部 | 腹神經節融入至腦部 | |
|---|---|---|
| 鱟 | 所有 | 只有首二節 |
| 蠍子 | 所有 | 沒有 |
| 古疣亞目 | 只有首二節 | 沒有 |
| 其他蛛形綱 | 所有 | 所有 |
螯肢亞門的神經系統都是基於節肢動物的一對標準神經索,每節的神經索上都有神經節。它們的腦部是神經節的融合,位於口器之後。[2]不過,由於螯肢二門的第一節會消失,所以它們的腦部只包含一對口前神經節,而不像其他昆蟲有兩對。[2]其他神經節融入至腦部的方式各有不同。鱟的腦部包含了所有頭胸部及首兩腹節的神經節,而其他腹節的神經節則保持分離。[2]除了蠍子以外,大部份現存蛛形綱的所有神經節都在頭胸部融合成單一團,而腹部則完全沒有神經節。[2]古疣亞目腹部及頭胸後部的神經節並沒有融合[9],而蠍子頭胸部的神經節融合在一起,但腹部的則保持分離。[2]
感覺
螯肢亞門的表皮阻隔了外界的資訊,只透過傳感器或神經系統的連接來獲得感覺。蜘蛛及其他節肢動物都改進了其表皮成一系列的傳感器。絨毛是觸覺及振動的傳感器,由強烈的接觸至非常弱的氣流也可以感知。化學傳感器提供了與味覺及嗅覺一樣的功用。[2]
現存螯肢亞門擁有複眼及單眼,複眼位於頭部兩側,單眼位於中央。鱟的眼睛可以在沒有形成影像的情況下偵測運動。[2]蠅虎科有非常闊的視覺[2],其主眼比蜻蜓的準確多十倍。[10]
繁殖

鱟是體外受精的,即精子及卵子是在體外相遇。它們的幼體像三葉蟲,只有兩對書鰓,在脫殼後會長多三對。[2]
蛛形綱是體內受精,一些會是直接的,即雄性及雌性的生殖器有直接接觸。另外也有一些受精是間接的,雄性會以它們鬚肢作為針筒將精子注入雌性的生殖口[2],或是雄性產出精囊,雌性將之放入體內。[2]示愛是十分普遍的,尤其是在強壯的掠食者,雄性要避免在交配前被雌性吃掉。大部份蛛形綱都是產卵的,而所有蠍子及一些蟎都會將卵藏在體內直至孵化出幼體。[2]
螯肢亞門的育嬰程度各有不同。蠍子會將幼體放在背上直至它們第一次脫殼,而一些物種就會一直留在母親身邊。[11]一些蜘蛛會照顧其幼體,如狼蛛科的幼體會抓著母親的絨毛[2],而母親亦會報以獵物作為回應。[12]
演化歷史
化石紀錄
螯肢亞門的化石紀錄有很大的斷層,這是由於牠們的外骨骼是有機的,很難作為化石被保存下來,除非是在特別適合保存軟組織的地層才能有所發現。伯吉斯頁岩中屬於5億500萬年前的多鬚蟲及西德尼蟲都被分類到螯肢亞門中,因為前者有體區分段,而後者則有像劍尾目的附肢。支序分類學分析卻認為它們有更多的特徵並不屬於螯肢亞門。5億1800萬年前的撫仙湖蟲是否屬於螯肢亞門激起了爭論。另一個寒武紀的科氏驚異蟲原先被分類到光楯目(Aglaspidida)中,但有可能是屬於板足鲎目。若這些任何一種是與螯肢亞門有關,化石斷層最少也有4300萬年。[13]
已知最古老的蛛形綱是Palaeotarbus jerami,源自4億2000萬年前的志留紀。牠們擁有三角形的頭胸部及有節的腹部,有八對腳及一對鬚肢。[14][15]
3億9300萬年前泥盆紀的Attercopus fimbriunguis是最早擁有生產絲器官的[16],但卻沒有噴絲板,故並非真正的蜘蛛。[17]有幾種石炭紀的蜘蛛都是屬於中突蛛亞目。[16]
志留紀晚期的黎蝎被分類為蠍子,但明顯與現今的蠍子有所分別。牠們是完全水生的,用鰓呼吸。其口器位於頭部底,接近第一對腳之間,有點像已滅絕的板足鲎目及現存的鱟。[18]4億200萬年前泥盆紀早期的陸生蠍子化石擁有書肺。[19]
系统发育树
- 螯肢亚门 Chelicerata
- 其他相關生物
- †光楯目 Aglaspidida
- †Cheloniellida
- †西德尼虫属 Sidneyia
與其他的關係
傳統上螯肢亞門與其他主要的節肢動物門較為疏遠。但近期的分子種系發生學及對節肢動物的神經系統在胚胎時期的發育研究,顯示螯肢亞門與多足綱較為接近。不過,研究結果只是針對現存的節肢動物,若結合已滅絕的動物(如三葉蟲),結果可能會重返傳統的觀點上。[20]
主要亞類
一般認同螯肢亞門包含了蛛形綱、劍尾目及板足鲎目。[21]已滅絕的Chasmataspidida是板足鲎目下的亞類。[21][22]海蜘蛛綱傳統上分類在螯肢亞門之內,但有些特徵顯示它們是演化自最古老的節肢動物。[23]
不過自19世紀,螯肢亞門演化樹內的關係就已經被受爭議。2002年綜合現今螯肢亞門的RNA特徵及現今和已滅絕的螯肢亞門的解剖特徵後,為低層亞類提供了可靠的結果,但在高階的亞類中並不穩定。[24]2007年的研究提供了右側的演化樹,但卻仍有一些不明處。[21]
蠍子的分類位置特別受到爭論。一些早期被認為是蠍子的化石,如志留紀晚期的Proscorpius卻有鰓和是水生的,口器的位置就像板足鲎目及鱟一樣。[18]這可以有三個選擇:將這些水生的化石分類到其他地方;或接受蠍子並非單系群,而是有水生及陸生的;[18]或是將蠍子分類到較為接近板足鲎目的位置,而疏於蜘蛛及其他蛛形纲[5],蠍子因而並非屬於蛛形纲,而蛛形纲亦非單系群。[18]
多樣性
雖然不及昆蟲,但螯肢亞門仍是最多樣化的動物之一,其下有超過77000個現存物種。[25]估計仍有13萬種蜘蛛及50萬種蟎未被描述。[26]雖然最早的螯肢亞門及現存的海蜘蛛及劍尾目都是水生及呼吸溶氧,其他的都是呼吸空氣的[25],除了一些蜘蛛可以將蜘蛛網織成潛水鐘的形狀進入水中生活。[27]大部份現存的螯肢亞門都是掠食者,主要掠食細小的無脊椎動物。其他很多都是寄生、植食性、食腐性及食碎屑性的。[2][25]
| 現存螯肢亞門的多樣性 | ||
| 類別 | 已描述物種[25] | 食性 |
|---|---|---|
| 海蜘蛛綱 | 1,300 | 肉食性[25] |
| 劍尾目 | 4 | 肉食性[25] |
| 蜘蛛目 | 45,000 | 肉食性;[25]1種植食性[6] |
| 蜱蟎亞綱 | 32,000 | 肉食性、寄生、植食性及食碎屑性[2][25] |
| 盲蛛目 | 6,300 | 肉食性、植食性及食碎屑性[2] |
| 拟蝎目 | 3,200 | 肉食性[28] |
| 蠍目 | 1,750 | 肉食性[2] |
| 避日目 | 1,200 | 肉食性、雜食性[2] |
| 裂盾目 | 180 | |
| 无鞭目 | 100 | |
| 有鞭目 | 90 | 肉食性[29] |
| 鬚腳目 | 60 | |
| 節腹目 | 60 | |
與人類的關係
_Lorryia_formosa_2_edit.jpg.webp)
以往美國原住民是會吃鱟,用其尾棘製作矛頭,殼用來舀水。鱟曾被作為牲畜食物,但因為發現會令牲畜肉質的味道變壞。鱟的血液含有鱟阿米巴樣細胞溶解物(Limulus Amebocyte Lysate)可以用作細菌存在的試劑,以及用來偵測腦膜炎及一些癌症。[30]
在柬埔寨[31]及委內瑞拉南部[32]會煮食捕鳥蛛科,並視為是一種美食。蜘蛛毒素對環境的污染較傳統的殺蟲劑為低,因只會對昆蟲(而不是脊椎動物)起作用。[33]蜘蛛毒素也正研究用作醫藥來治療心律不整[34]、阿茲海默病[35]、中風[36]及勃起功能障礙。[37]由於蜘蛛絲輕巧而堅韌,基因工程師嘗試利用羊奶及葉子來製作蜘蛛絲。[38][39]於20世紀就有約100宗被蜘蛛咬死的個案[40],相對於水母刺死的1500宗為小。[41]
蠍子刺螯亦是相當危險的,例如在墨西哥每年就約有1000宗死亡個案,但在發展較先進的美國每年就只有1宗。大部份個案都是因人類誤闖蠍子巢穴而發生的。[42]蠍子毒素也正被開發來作為醫藥用途,包括治療腦癌及骨骼病症。[43][44]
蜱是寄生的,一些會傳播引發疾病的微生物及寄生蟲,一些的口水更會直接引起蜱癱瘓。[45]
一些蟎也會侵擾人類,它們咬或鑽入皮膚下而引起痕癢。那些侵擾齧齒目等動物的物種在失去主體時也會影響人類。[46]有三種蟎會對蜜蜂造成危害,其中一種的瓦蟎是養蜂人的勁敵。[47]蟎會引起多種過敏症,包括過敏性鼻炎、哮喘及濕疹,與及加劇異位性皮膚炎。[48]它們也是害蟲,但掠食性的蟎可以控制它們。[25][49]
參考
- Margulis, Lynn; Schwartz, Karlene. third. W.H. Freeman and Company. 1998. ISBN 0716730278.
- Ruppert, E.E., Fox, R.S., and Barnes, R.D. 7. Brooks/Cole. 2004: 518–84. ISBN 0030259827.
- Willmer, P., and Willmer, P.G. . Cambridge University Press. 1990: 275 [2008-10-14]. ISBN 0521337127.
- Braddy, S.J., Markus Poschmann, M., and Tetlie, O.E. . Biology Letters. 2008, 4 (1): 106–9. PMC 2412931
 . PMID 18029297. doi:10.1098/rsbl.2007.0491.
. PMID 18029297. doi:10.1098/rsbl.2007.0491. - Braddy, S.J., Aldridge, R.J., Gabbott, S.E., and Theron, J.N. . Lethaia. 1999, 32 (1): 72–4.
- Meehan, C,J. Olson, E.J. and Curry, R.L. . 93rd ESA Annual Meeting. 21 August 2008 [2008-10-10]. (原始内容存档于2019-12-01).
- Jackson, R.R; et al. (PDF). J. Zool. Lond. 2001, 255: 25–9 [2010-09-11]. doi:10.1017/S095283690100108X. (原始内容存档 (PDF)于2009-03-18).
- Gould, S.J. . Hutchinson Radius. 1990: 105. ISBN 0091742714.
- Coddington, J.A. & Levi, H.W. . Annu. Rev. Ecol. Syst. 1991, 22: 565–92. doi:10.1146/annurev.es.22.110191.003025.
- Harland, D.P., and Jackson, R.R. (PDF). Cimbebasia. 2000, 16: 231–240 [2008-10-11]. (原始内容 (PDF)存档于2006-09-28).
- Lourenço, W.R. . Toft, S., and Scharff, N. (编). (PDF). Aarhus University Press. 2002: 71–85 [2008-09-28]. ISBN 87-7934-001-6. (原始内容存档 (PDF)于2008-10-03).
- Foelix, R.F. . . Oxford University Press US. 1996: 176–212 [2008-10-08]. ISBN 0195095944.
- Wills, M.A. . Geological Journal. 2001, 36 (3-4): 187–210. doi:10.1002/gj.882.
- Dunlop, J.A. (PDF). Palaeontology. September 1996, 39 (3): 605–14 [2008-10-12]. (原始内容存档 (PDF)于2008-12-16).
- Dunlop, J.A. . Palaeontology: 191.
- Vollrath, F., and Selden, P.A. (PDF). Annual Review of Ecology, Evolution, and Systematics. December 2007, 38: 819–46 [2008-10-12]. doi:10.1146/annurev.ecolsys.37.091305.110221. (原始内容 (PDF)存档于2008-12-09).
- Selden, P.A., and Shear, W.A. (PDF). Nature Precedings. July 2008 [2008-10-12]. (原始内容存档 (PDF)于2016-08-10).
- Weygoldt, P. . Experimental and Applied Acarology. February 1998, 22 (2): 63–79. doi:10.1023/A:1006037525704.
- Shear, W.A., Gensel, P.G. and Jeram, A.J. . Nature. December 1996, 384: 555–7. doi:10.1038/384555a0.
- Jenner, R.A. . Integrative and Comparative Biology. 2006, 46 (2): 93–103. doi:10.1093/icb/icj014.
- Schultz, J.W. . Zoological Journal of the Linnean Society. 2007, 150: 221–65. doi:10.1111/j.1096-3642.2007.00284.x.
- O. Tetlie, E., and Braddy, S.J. . Transactions of the Royal Society of Edinburgh: Earth Sciences. 2003, 94: 227–34. doi:10.1017/S0263593300000638.
- Poschmann, M., and Dunlop, J.A. . Palaeontology. 2006, 49 (5): 983–9. doi:10.1111/j.1475-4983.2006.00583.x.
- Gonzalo Giribet G., Edgecombe, G.D., Wheeler, W.C., and Babbitt, C. . Cladistics. 2002, 18 (1): 5–70. PMID 14552352. doi:10.1111/j.1096-0031.2002.tb00140.x.
- Shultz, J.W., , , John Wiley & Sons, Ltd., 2001, doi:10.1038/npg.els.0001605
- (PDF), Department of the Environment and Heritage, Australian Government, September 2005 [2010-03-29], (原始内容存档于2007-09-11)
- Schütz, D., and Taborsky, M. (PDF). Evolutionary Ecology Research. 2003, 5 (1): 105–17 [2008-10-11]. (原始内容 (PDF)存档于2008-12-16).
- . Pennsylvania State University. [2008-10-26]. (原始内容存档于2008-07-04).
- Harvey, M.S. (PDF). Journal of Arachnology. 2002, 30: 357–72 [2008-10-26]. (原始内容 (PDF)存档于2010-12-13).
- Heard, W. (PDF). University of South Florida. [2008-08-25]. (原始内容 (PDF)存档于2017-02-19).
- Ray, N. . LonelyPlanet Publications. 2002: 308. ISBN 1-74059-111-9.
- Weil, C. . Plume. 2006 [2008-10-03]. ISBN 0452287006. (原始内容存档于2011-05-11).
- . National Science Foundation (USA). [2008-10-11]. (原始内容存档于2012-06-02).
- Novak, K. . Nature Medicine. 2001, 7 (155): 155. PMID 11175840. doi:10.1038/84588.
- Lewis, R.J., and Garcia, M.L. (PDF). Nature Reviews Drug Discovery. October 2003, 2 (10): 790–802 [2008-10-11]. PMID 14526382. doi:10.1038/nrd1197. (原始内容 (PDF)存档于2008-12-16).
- Bogin, O. (PDF). Modulator. Spring 2005, (19) [2008-10-11]. (原始内容 (PDF)存档于2008-12-09).
- Andrade, E., Villanova, F., Borra, P.; et al. . British Journal of Urology International. June 2008, 102 (7): 835–7. PMID 18537953. doi:10.1111/j.1464-410X.2008.07762.x.
- Hinman, M.B., Jones J.A., and Lewis, R.W. (PDF). Trends in Biotechnology. September 2000, 18 (9): 374–9 [2008-10-19]. PMID 10942961. doi:10.1016/S0167-7799(00)01481-5. (原始内容 (PDF)存档于2008-12-16).
- Menassa, R., Zhu, H., Karatzas, C.N., Lazaris, A., Richman, A., and Brandle, J. . Plant Biotechnology Journal. June 2004, 2 (5): 431–8. PMID 17168889. doi:10.1111/j.1467-7652.2004.00087.x.
- Diaz, J.H. . American Journal of Tropical Medicine and Hygiene. August 1, 2004, 71 (2): 239–50 [2008-10-11]. PMID 15306718. (原始内容存档于2011-05-11).
- Williamson, J.A., Fenner, P.J., Burnett, J.W., and Rifkin, J. . UNSW Press. 1996: 65–8 [2008-10-03]. ISBN 0868402796.
- Cheng, D., Dattaro, J.A., and Yakobi, R. . WebMD. [2008-10-25]. (原始内容存档于2008-12-06).
- . BBC News. 2006-07-30 [2008-10-25]. (原始内容存档于2019-08-14).
- . Harvard University. [2008-10-25]. (原始内容存档于2010-06-20).
- Goodman, Jesse L.; Dennis, David Tappen; Sonenshine, Daniel E. . ASM Press. 2005: 114–401 [2010-03-29]. ISBN 9781555812386. (原始内容存档于2012-10-26).
- Potter, M.F. . University of Kentucky College of Agriculture. [2008-10-25]. (原始内容存档于2009-01-08).
- Jong, D.D., Morse, R.A., and Eickwort, G.C. . Annual Review of Entomology. January 1982, 27: 229–52. doi:10.1146/annurev.en.27.010182.001305.
- Klenerman, Paul; Lipworth, Brian; authors. . NetDoctor. [2008-02-20]. (原始内容存档于2008-02-11).
- Osakabe, M. . Experimental & applied acarology. 2002, 26 (3-4): 219–30. doi:10.1023/A:1021116121604.