雙冠龍屬
雙冠龍(屬名:,發音:/daɪˌloʊfəˈsɔːrəs, -foʊ-/[1] dy-LOHF-o-SOR-əs,或譯雙脊龍)是種早期的中大型肉食性獸腳類恐龍,生存於約1億9300萬年前早侏羅世錫內穆階的北美洲。1940年在美國亞利桑那州北部發現三個骨骼,其中保存最好的兩個採集於1942年。最完整的標本成為正模標本,於1954年由塞繆爾·保羅·威爾斯命名為斑龍屬(當時是個分類垃圾桶)的新種魏氏斑龍(Megalosaurus wetherilli)。1964年威爾斯發現一個較大的骨骼也屬於同一物種,並意識到這種動物的頭骨其實長有頭冠,於是在命名時隔十幾年後的1970年建立為獨立的雙冠龍屬,模式種為魏氏雙冠龍(Dilophosaurus wetherilli),屬名在古希臘語意為「兩個脊冠的蜥蜴」,種名紀念一位納瓦霍人議員約翰·魏瑟瑞(John Wetherill)。1984年威爾斯發表了關於雙冠龍的詳細骨骼描述。後續還進一步發現更多標本,其中包括一具幼體。一些足跡化石也被歸入雙冠龍,其中還有休息留下的痕跡。一隻於1984年在中國雲南祿豐組發現近乎完整的標本於1993年被命名為中國雙冠龍(Dilophosaurus sinensis),後來發現其實屬於中國龍屬。2020年亞當·馬許(Adam D. Marsh)和提摩西·羅威(Timothy B. Rowe)對雙冠龍的所有已知標本進行了全面性的重新描述,這項耗時7年的研究澄清了許多過去在研究雙冠龍時所遇到的問題。
| 雙冠龍屬 | |
|---|---|
 | |
| 正模標本UCMP 37302的骨架模型,呈現被掩埋的原況姿勢,展示於皇家安大略博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 獸腳亞目 Theropoda |
| 科: | †雙冠龍科 Dilophosauridae |
| 属: | †雙冠龍屬 Dilophosaurus Welles, 1970 |
| 模式種 | |
| †魏氏雙冠龍 Dilophosaurus wetherilli (Welles, 1954) | |
| 異名 | |
| |
雙冠龍身長約7公尺,體重約400公斤,是年代最早期的大型掠食性獸腳類之一,也是已知當時北美洲體型最大的陸生動物。雙冠龍身型纖細且結構輕盈,頭骨特別大但纖巧。吻部狹窄,上頜鼻骨下方有個凹槽。頭骨上方有一對縱向的拱形嵴冠,完整的形狀尚未確認,但相信生前可能覆蓋著角質層而更大。下頜前部纖細、後部深。牙齒長、彎曲、薄且側扁。下頜的牙齒比上頜的小許多。大多數牙齒前後緣都有鋸齒狀。頸部很長,脊椎中空所以非常輕巧。手臂強壯,上臂骨長而纖細。手部有四根手指;第一指短而強壯,長有大型指爪;第二指較長且細,爪子較小;第四指是退化的痕跡器官。股骨大,腳很粗壯,並長著大趾爪。
雙冠龍與近緣的龍獵龍構成雙冠龍科,演化位置介於腔骨龍科與後來更進階的堅尾龍類之間。雙冠龍以雙足行走,可能是活躍主動的掠食者,並能捕殺大型獵物,也可能以較小型動物或魚類為食。因為前肢短小且活動範圍有限,可能主要透過嘴部來襲擊獵物。頭冠確切功能尚未知曉,但相信對於打鬥來說過於脆弱,較可能用作視覺展示用途,如種內識別或性選擇。成長過程可能十分迅速,生命階段早期每年增長程度達到30至35公斤。正模標本的個體顯示出許多古病理跡象,包括發育異常及損傷後的癒合痕跡。雙冠龍的化石來自凱恩塔層,與巨歿龍、莎拉龍共享棲地。雙冠龍曾在侏羅紀公園及其改編電影中登場,當中賦予其噴射毒液及展開頸邊皮摺的虛構能力,且只有狗一般的大小,遠遠小於真實動物的體型。根據在康乃狄克州發現的足跡,雙冠龍被指定為該州的州恐龍。
發現及研究歷史

1942年夏天,古生物學家查爾斯·坎普帶領來自加州大學古生物博物館的團隊前往北亞利桑那納瓦霍縣找尋脊椎動物化石。此消息於當地原住民間傳播開來,納瓦霍的傑西·威廉斯(Jesse Williams)帶領三位團隊成員去到他於1940年發現的一些骨骸。當地屬於凱恩塔層的一部分,位於納瓦荷保留地圖巴市附近卡梅倫以北32公里處。在紫色頁岩中發現了三具恐龍骨骼,排列成三角形,一側長約9.1公尺。第一具骨骼近乎完整,僅缺乏頭骨前部、部分骨盆與一些脊椎。第二具骨骼遭到嚴重侵蝕,包括頭骨前部、下頜、一些脊椎、肢骨、一隻關節連接的手。第三具骨骼被侵蝕到只剩下脊椎碎片。第一具完好的骨骼經歷10個工作天後以石膏包裹起來並運上卡車載走。第二具骨骼採集十分容易,因為已被風化到完全露出地表。但遺憾的是,第三具骨骼幾乎徹底遺失了。[2][3][4]
第一具近乎完整的標本被送至加州大學古生物博物館進行清修工作,在古生物學家浣·蘭斯頓的監督下,過程由三個人進行並耗時兩年。該骨架以淺浮雕的樣貌裝設於牆面上,尾部上彎、頸部伸直、左腿上移以提升能見度,而骨架其餘位置則維持在埋藏原位。由於骨架本身的頭骨已碎裂,重建過程參考了第一具標本的頭骨後部與第二具標本的頭骨前部。骨盆參考異特龍來進行重建。而腳部亦是重建。雖然完整度還不是很高,卻已是當時保存最良好的獸腳類化石之一。1954年,曾參與挖掘工作的其中一名古生物學家塞繆爾·保羅·威爾斯將這種恐龍初步描述為當時現有的斑龍屬底下的一個新物種:魏氏斑龍(M. wetherilli)。近乎完整的第一具標本UCMP 37302被指定為該物種的正模標本,而第二具標本UCMP 37303則作為副模標本。種名紀念納瓦霍議員約翰·魏瑟瑞(John Wetherill),威爾斯稱其為「探險家、科學家之友及值得信賴的商人」。魏瑟瑞的姪子米爾頓首先告知了化石考察隊。威爾斯將這個新物種歸入斑龍是基於其與巴氏斑龍(M. bucklandii)的肢體比例相近,且他沒有發現兩者之間有太大的差異。在當時斑龍屬是一個分類垃圾桶,其中丟進了許多未知、不限年代及地點的獸腳類。[2][5][3][6]
威爾斯於1964年回到圖巴市確認凱恩塔層的年代(曾被認為是晚三疊世,但威爾斯認為是早至中侏羅世),並在1942年那批骨骼以南約400公尺處發現了另一具骨骼。在北亞利桑那博物館的威廉·比德(William Breed)等人的協助下,採集了近乎完整的UCMP 77270標本。在清修該標本的過程中意識到這是一隻更大的魏氏斑龍個體,且頭骨頂部有兩個頭冠。因為是塊骨頭薄片,其中一片頭冠起初被認為屬於頭骨缺失的左側部分(已被食腐動物從原位置挖出)。當意識到這明顯是個頭冠時,基於右頭冠位於中線右側,且延中線長呈凹陷,而認為另一片對應的頭冠應位於左側。此發現導致重新對正模標本進行檢驗,並發現該標本有兩根向上延伸的細骨基部已被擠壓成一坨。這也屬於頭冠一部分,但過去被誤認為屬於位移的顴骨。1942年的兩具標本被認為是年輕個體,而1964年的標本是個成年個體,體型大於其他標本約1/3。[2][7][8]威爾斯後來回憶道:這個頭冠就像「在蠕蟲身上發現翅膀」般令人出乎意料。[9]

.jpg.webp)
威爾斯和一名助理隨後根據新標本對正模標本的模型進行修正,包括復原頭冠、重作骨盆、加長頸肋及使彼此更靠近。在研究各種北美及歐洲的獸腳類骨骼後,威爾斯意識到這個物種不屬於斑龍,而需要一個新的屬名。在當時還未曾見過其他頭上長有大頭冠的獸腳類恐龍,於是引起了古生物學界的關注。製作了一個正模標本的模型,並將玻璃纖維材質的複製品提供於各種展示;為了易於標示展示品,威爾斯決定先在一篇簡略的聲明中命名新屬,而非拖到詳細描述論文發佈的時候。1970年威爾斯建立了新屬名雙冠龍,由古希臘語的δι/di(雙)、λόφος/lophos(嵴冠)、σαυρος/sauros(蜥蜴)組成:全意為「兩個頭冠的蜥蜴」。1984年,威爾斯發表了對雙冠龍的詳細骨學描述,但沒有涵蓋1964年的標本,因為當時他認為這是另一種不同的動物。[2][7][11][8][10]雙冠龍是第一種知名的侏羅紀早期獸腳類,且依舊是那個時代保存最良好的例子之一。[5]
2001年古生物學家羅伯·給在北亞利桑那博物館的館藏中鑑定出至少三具新的雙冠龍標本(基於三個恥骨碎片及兩個不同尺寸的股骨而推估出來的數量)。這些標本是於1978年在距離初始標本發現地約190公里處的岩頭矩(Rock Head Quadrangle)所採集,並被標示為「大型獸腳類」的名下。雖然多數材料已毀損,但其中涵蓋一些早期標本中缺失的部位(包括部分骨盆及幾根肋骨)而具有重要意義。這些館藏的其中一些材料來自幼體(MNA P1.3181),是雙冠龍已知最年幼的標本,也是北美已知最古老的獸腳類幼體之一,僅稍晚於某些腔骨龍標本。右體標本包括部分肱骨、部分腓骨、牙齒碎片。[12]2005年古生物學家羅納·泰可斯基(Ronald S. Tykoski)將一具來自亞利桑那金泉的TMM 43646-140標本歸入雙冠龍,但2012年古生物學家馬修·卡拉諾(Matthew T. Carrano)與同事發現某些細節處有所差異。[13][14]
2020年古生物學家亞當·馬許(Adam D. Marsh)和提摩西·羅威(Timothy B. Rowe)針對現時已知所有的雙冠龍標本發表了全面的重新描述,其中包括自1964年以來長期未曾被描述的UCMP 77270標本,同時也排除了一些過去曾歸入、過於破碎而難以鑑別的標本,且還透過傑西·威廉斯親戚的協助下重新定位了化石的初始發現地。[6][15]在一次訪談中馬許稱雙冠龍是「所知最少的知名恐龍」("best worst-known dinosaur"),因為雖然這種恐龍早在80年前就被發現了,人們對牠的了解卻一直以來都很模糊。其中一個主要的問題在於,過去對標本的研究都未指明哪些是原本的化石、哪些是石膏重建的部分,而後來的研究人員皆參考威爾斯1984年的論文進行後續研究,造成理解雙冠龍解剖結構上的混淆。馬許費時七年來研究這些標本,以為了澄清各種關於雙冠龍的疑問,其中也包括由他的指導教授羅威於20年前發現的兩個標本。[16]
過去歸入種

1984年威爾斯表示,於1964年發現的UCMP 77270標本不屬於雙冠龍,根據頭骨、脊椎、股骨上的差異,而判定屬於一個未知的新屬。他維持原有的觀點,即兩屬都具有頭冠,但雙冠龍的確切頭冠形狀是未知的。[2]威爾斯於1997年逝世,當時他還沒能來得及命名這隻所謂的新屬恐龍,此後這些標本存在不同屬的觀點普遍受到後世的忽視或遺忘。[5]1999年業餘古生物學家史蒂芬·皮克林(Stephan Pickering)在私人著作中將1964年標本發表為雙冠龍的新種比氏雙冠龍(Dilophosaurus "breedorum"),種名紀念對採集標本有貢獻的威廉·比德(William Breed)。這個物種被視為裸名(非官方正式發表的無效學名)。而2005年給的研究指出比氏種與其他魏氏種的標本沒有任何顯著的差異。[17][18]2012年,卡拉諾與同事發現1964年標本與正模標本間的差異,但認為這只是個體上自然存在的變異、而非種間差異。[13]2014年,古生物學家奎斯妥夫·罕垂克斯(Christophe Hendrickx)和奧塔維歐·馬諦烏斯(Octávio Mateus)表示,依照標本存在不同的頭骨特徵、及彼此出土地層的差異,已知標本仍可能屬於兩個不同物種,但仍需待對這些標本進行進一步的描述。[19]2020年,馬許和羅威的研究得出結論:在所有已知的雙冠龍標本中僅存在單一物種魏氏雙冠龍,而彼此間的形態差異是受到成熟度及保存程度上的不同所影響;另外他們也發現標本出土地層並沒有特別大的差距。[6]
1987年,在中國雲南省祿豐組發現了一具近乎完整的獸腳類骨骼,編號KMV 8701標本。整體外形近似雙冠龍,有一對頭冠、前上頜骨與上頜骨間有道間隙,但在一些細節上與雙冠龍有所差異。1993年古生物學家胡紹錦將之命名為雙冠龍的新種:中國雙冠龍(D. sinensis,當時舊譯為中國雙脊龍)。[20]1998年古生物學家馬修·拉曼納和同事發現中國雙冠龍與另一種來自同一地層、於1940年命名的三疊中國龍(Sinosaurus triassicus)幾乎一致。[21]2013年邢立達和同事確認了前述論點,而王國富和同事於2017年認同將中國種納入中國龍屬下,卻表示這個物種可能與三疊種不同,而維持將中國中國龍(S. sinensis)視為獨立種。[22][23]
生痕化石

已有數個根據痕跡化石建立的遺跡分類單元被認為屬於雙冠龍或類似的獸腳類。1971年威爾斯記載了來自北亞利桑納凱恩塔層的恐龍足跡,位於初始雙冠龍骨骸之下14公尺及112公尺的兩層。較下方的足跡為三趾型,可能由雙冠龍所留下;威爾斯根據其建立了新的足跡分類單元威氏雙冠龍足跡(Dilophosauripus williamsi),種名紀念第一具雙冠龍骨骸的發現者威廉斯。其正模標本是塊大型腳印模型UCMP 79690-4,其餘參考標本有另外三個腳印模型。[24]1984年威爾斯表示並未發現任何方法足以證明或反駁這些痕跡屬於雙冠龍。[2]1996年邁可·莫拉萊斯(Michael Morales)、史考特·伯奇(Scott Bulkey)記載了一條來自凱恩塔層遺跡分類單元真雷龍足跡,由某種非常大型的獸腳類所留下。他們註記道這可能是由一個非常大型的雙冠龍個體所製造,但又不太合理,因為依照痕跡估計製造者的臀高高達2.83至2.99公尺,明顯高於雙冠龍的1.5至1.75公尺。[25]

傑拉德·哲林斯基(Gerard Gierliński)檢驗了來自波蘭聖十字山脈的三趾足跡,並於1991年表示屬於某種類似雙冠龍的獸腳類。他據此命名了新的足跡分類單元薩提柯夫蹺行足跡(Grallator (Eubrontes) soltykovensis),並指定MGIW 1560.11.12足模為正模標本。[26]1994年哲林斯基還將1974年在瑞士霍加納斯層發現的足跡歸入薩提柯夫蹺行足跡。[27]1996年哲林斯基記載了來自麻塞諸塞州透納瀑布層的AC 1/7痕跡,是一道休息痕跡,他相信其中帶有羽毛印痕,並屬於某種類似雙冠龍或理理恩龍的獸腳類,歸入微型蹺行足跡(Grallator minisculus)。[28]2004年馬汀·昆札特(Martin Kundrát)認同該痕跡顯示出羽毛印痕,但這番詮釋分別於2003年馬汀·洛克里及2004年安東尼·傑馬汀的研究中受到質疑,他們認為這些痕跡是移動時產生的碎屑沉積物。馬汀等人還將痕跡歸入Fulicopus lyellii。[29][30][31]
2003年羅伯·偉因斯(Robert E. Weems)提出真雷龍足跡並非由獸腳類製造,而是由類似板龍的蜥腳形亞目,從而排除了雙冠龍的可能性。偉因斯認為由威爾斯於1971年命名的霍皮凱恩塔足跡(Kayentapus hopii)才與雙冠龍最為吻合。[32]歸入雙冠龍的主要依據來自足跡第三和第四趾間的角度,符合正模標本觀察到相似的第四趾開展。同樣於2003年,艾瑪·蕊茵芙(Emma Rainforth)反駁道雙冠龍正模標本腳趾的開展僅是化石變形的結果,真雷龍足跡依然與雙冠龍相符。[33][34]2006年史賓賽·盧卡斯和同事表示,真雷龍足跡是由類似雙冠龍的獸腳類所製造已受到普遍的共識,從而反對偉因斯的觀點。[35]
2006年偉因斯為其2003年對真雷龍足跡的論點提出了辯駁,並表示維吉尼亞州寇沛珀採石場(Culpeper Quarry)的數個凱恩塔足跡可能是由類似雙冠龍的動物所製造的。他推測一些與足跡關聯的圓形印痕,可能代表著手指沒印上去的手部痕跡,將之解釋成四足行動的證據。[34]2009年米爾納等人使用新的命名組合薩提柯夫凱恩塔足跡(Kayentapus soltykovensis),並表示雙冠龍足跡、真雷龍足跡、凱恩塔足跡之間可能沒有明顯的差異。他們認為用以界定雙冠龍足跡的長趾爪印可能由拖曳所造成,並發現巨雙足跡(Gigandipus)與近蜥龍足跡(Anchisauripus)也可能同樣是真雷龍足跡的變異形態。他們指出足跡物種間的差異可能是由足跡製造者與底質交互作用的結果,而非分類學上的差異。他們還發現在猶他州繆內孚層聖喬治恐龍發現遺址,雖然該地區沒有發現過雙冠龍骨骼,且年代稍比凱恩塔層要古老一些,但當地的真雷龍足跡與休息痕跡(編號SGDS 18.T1)卻與雙冠龍相符合。[36]2019年偉因斯表示真雷龍足跡並非雙冠龍纖細的腳,而是由雙足蜥腳形亞目近蜥龍所製造的。[37]
描述

雙冠龍是年代最早期的大型掠食性恐龍之一,體型在獸腳類中屬於中間值,但仍較其他晚期獸腳類小型。[2][5]牠同時也是侏羅紀早期北美洲已知最大的陸生動物。[6]其身體纖細、結構輕盈,體型堪比現代棕熊。[5][38][39]已知最大隻的標本個體體重約400公斤、身長測量有7公尺、頭骨長59公分。較小的正模標本個體體重約283公斤、6.03公尺長、臀高約1.36公尺、頭骨長52.3公分。[38][40]一個由類似雙冠龍或理理恩龍的獸腳類所遺留下的休息痕跡被某些研究人員描述成在腹部與腳部周圍顯示出羽毛印痕,結構類似絨羽。[28][29]其他研究人員則將這類印痕解釋為恐龍移動時產生的碎屑沉積物,但仍不排除痕跡製造者本身長有羽毛的可能性。[30][31]
頭骨






與全身骨骼相比,雙冠龍的頭骨比例較大,但結構精緻。吻部前視狹窄,往圓潤尖端越變越窄。前上頜骨側視長而低矮,前方呈球莖狀,其外表面從吻部向鼻孔凸度趨緩。與多數獸腳類相比,鼻孔位置更後方。前上頜骨彼此關節緊密相連,前上頜骨僅在上頜中部與上頜骨連接,側面未有相交,彼此透過骨骼前後緊鎖的突起構成堅固的關節。前上頜骨後下方與上頜骨之間構成一道壁,稱為扭結的鼻下縫(subnarial gap),這也存在於腔骨龍科等恐龍身上。次鼻縫構成齒列中的齒間隙(diastema)。鼻下縫內坐落前上頜骨齒列後面的深凹,稱為鼻下凹(subnarial pit),由前上頜骨向下的骨突圍成壁。[2][38][41][7][6]
前上頜骨外表面覆蓋著大小不等的孔洞。前上頜骨兩條後向延伸突起的上部長而低矮,構成加長鼻孔的大部分上緣,前傾使底部輪廓呈凹狀。前上頜骨下側包括牙槽呈橢圓形。上頜骨淺,在眶前孔周圍凹陷,構成一前圓凹陷,比上頜骨其餘部分還光滑。上眶前孔於前彎處開放至此凹陷。上頜骨側邊、牙槽上方有大孔洞。從鼻下凹延至上頜骨齒間板基部有道後向的深營養孔。[2]

雙冠龍頭骨頂端水平向有一對高而薄、呈拱形或板狀的嵴冠。頭冠/鼻淚冠(nasolacrimal crests)始於前上頜骨上的低矮稜脊,主要由鼻骨及淚骨的向上擴張所構成。這些骨頭在發育時彼此融合成一塊,而無法分辨出其間的縫線。淚骨擴張成厚、有皺紋的眶前凸部,於眼眶上緣構成弧形,支撐頭冠後方的底部。雙冠龍的獨特之處在於,眼眶上緣繼續延伸向後,止於眼眶後方一道接近三角形、略外彎的小突起。由於該突起上表面僅一小部分保存完整,推測頭冠其餘部分可能在頭骨上隆起超過12毫米。UCMP 77270保存的頭冠部分在眶前孔長邊的中點附近處達到最高點。UCMP 77270保存了頭冠底部間的凹板,正視其朝上及兩側成80度角伸展開來。威爾斯覺得這樣的頭冠令人聯想成兩個頭冠的食火雞,而馬許和羅威表示頭冠生前可能覆蓋著角蛋白或角質化皮膚。他們指出,與有頭冠的珠雞相較之下,雙冠龍頭冠的角蛋白可能使本身尺寸比骨頭留下的部分要擴大更多。有鑑於只有一具標本的頭冠有大部分被保存下來,關於個體間頭冠是否存在差異,目前尚無法得知。[6][2][7][11][5][13]斷層掃描顯示氣囊(為骨骼提供強度並減輕重量的空腔)存在於圍繞腦殼的骨骼中,並連接到頭骨前部的竇腔。眶前孔與頭冠側邊接續,代表頭冠亦存在氣囊(至於其他多數獸腳類則是一道骨嵴構成眶前孔的上蓋)。[15]

眼眶呈橢圓形,往底部變窄。顴骨有兩個朝上的突起,第一個突起構成眶前孔、眼眶下緣的一部分。方骨至外顳孔(眼眶後方的洞)的突起構成了腎形輪廓。枕骨大孔(腦殼後方的大孔洞)寬度約為枕髁的一半。枕髁呈心形,頸短、側面有凹陷。[2]下頜骨前端纖巧,但與頭骨相連的關節區龐大。下頜孔周圍區域很深。與腔骨龍相比雙冠龍的下頜孔很小,由前向後縮減,為本屬獨有特徵。齒骨形成一道上彎的下巴,而非尖利。下巴尖端有個大孔,一排小孔大致平行於齒骨上緣。內側下頜兩半連接處平坦光滑,未顯示出彼此融合的跡象。梅氏孔沿齒骨外側延伸。隅骨側面與方骨關節前有個獨特的錐形突,該橫脊構成一隔板。下頜後關節突長。[2][41][6]
雙冠龍每個前上頜骨有4顆牙、每個上頜骨有12顆牙、每個齒骨有18顆牙。牙齒普遍長、薄且下彎,基部相對小。牙齒側扁,底部橫切面呈橢圓形、上視呈透鏡狀、內外側略凹。上頜骨最大牙位於第四牙槽內或附近,牙冠高度朝後降低。上頜骨第一牙由其牙槽略向前,因前上頜突下緣上翹所致。齒骨的牙齒比上頜骨的小許多。雙冠龍與某些腔骨龍科的齒骨第三或第四牙最大,而似乎與上頜鼻下凹處吻合。多數牙齒前後緣有鋸齒狀,與縱凹相抵,前面較小。前緣約31至41鋸、後緣約29至33鋸。至少前上頜骨第二及第三牙有鋸齒,但第四牙沒有。牙齒表面覆蓋一層薄琺瑯質,厚約0.1至0.15公厘,朝基部延伸很遠。牙槽為橢圓到近乎圓形,並大於其所包含牙齒的基部,因此在嘴內可能固定得鬆散。雖然牙槽數顯示牙齒可能排列擁擠,但牙槽較大而牙齒彼此距離較遠。頜骨內含有不同階段的替生齒。牙齒間的齒間板非常低矮。[2][41][12]
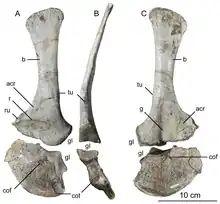
顱後骨骼






雙冠龍有10節頸椎、14節背椎、45節尾椎,氣囊生長其中。牠的頸部很長,從頸部到頭部間呈彎曲近90度角,使頭骨維持水平姿勢。頸椎不尋常地輕,其椎體(centra)由側凹(pleurocoels)與內凹(centrocoels)所鑿成空心。頸椎弓也有氣孔(pneumatic fossae/chonoses),其圓錐形凹陷之大使分隔骨有時薄如紙張。椎體呈平-凹型,即前部平坦或微凸、後部凹入,類似角鼻龍。這代表著就算頸肋長而重疊並與椎體融合,頸部依然高度靈活。[2][41][6][15]
寰椎(連接頭骨的第一頸椎)有小方形椎體,前端凹面,在頭骨後部構成一連接枕髁的杯狀。樞椎(第二頸椎)厚重,其後關節突與第三頸椎上彎的前關節突相連。椎體及神經棘長而低矮,側視呈階梯狀,前後構成「肩部」、中央有較高的「帽部」,使俯視呈馬爾他十字形,是雙冠龍的獨有特徵。後部中央椎板出現連續變化,延頸部分叉與結合,是為獨有特徵。背椎神經棘也低矮、前後擴張,構成韌帶牢固附著處。額外椎板由中部背椎的前中央椎板與後中央椎板伸出,是為獨有特徵。佔據腸骨長邊的薦椎似乎未融合。第一薦椎的肋骨與腸骨的前髖臼突相連,是為獨有特徵。尾椎椎體長度相當一致,但直徑朝後方漸小,橫切面由橢圓形變為圓形。[2][41][6]






肩胛骨長度中等,內側隨身體曲線而凹彎。肩胛骨寬闊,尤其上部,呈矩形,是為獨有特徵。鳥喙骨呈橢圓形,未與肩胛骨融合。鳥喙骨後下部二頭肌結節旁有個橫壁,是為獨有特徵。手臂強有力,有深凹陷及厚實突起,作為肌肉及韌帶的附著。肱骨大而纖細,尺骨粗而筆直,肘突厚實。手部有四根手指:第一指較後兩指短但強壯,配有大型指爪;後兩指長而纖細,爪小;爪子彎曲而鋒利;第四指退化成無功能的痕跡器官。[2][41][6]
腸骨脊在腸骨體柄(下突)處達到最高,外側呈凹形。恥骨靴僅略為擴張,而坐骨下端擴張較大,其軸也非常纖細。後肢龐大,股骨略長於脛骨,與腔骨龍之類的相反。股骨巨大,軸彎曲呈S形,大轉子位於軸中央。脛骨結節下端擴張並發育良好。距骨由脛骨與跟骨間分離出來,佔據腓骨窩的一半。腳部長而粗壯,三個發達腳趾帶有大爪,爪子彎曲程度比手爪小得多。第三趾最為粗壯,第一趾抬高遠離地面。[2][41][42][6]
分類

威爾斯最初於1954年認為這是一種斑龍類,但1970年當頭冠被發現後他隨即修正了觀點。[7][3]到了1974年,威爾斯和羅伯·艾隆(Robert A. Long)認為雙冠龍是種角鼻龍下目。[43]1984年威爾斯發現雙冠龍同時具有虛骨龍類和肉食龍下目(進階型獸腳類的兩大分支)的特徵,並根據體型藉此提出這種分法是不準確的觀點。他發現雙冠龍與通常被歸類為敏捷龍科(Halticosauridae)的獸腳類最近緣,尤其是理理恩龍。[2]
1988年葛瑞格利·保羅將敏捷龍科列入腔骨龍科的分支,並認為雙冠龍是腔骨龍的直系後裔。保羅還認為棘龍科是倖存至晚期的雙冠龍,是基於兩者類似的吻部凹槽、鼻孔位置,以及重爪龍的細齒來推定。[38]1994年湯瑪斯·霍茨將雙冠龍與腔骨龍科一同置於腔骨龍超科,但彼此各為獨立分支。[44]2000年詹姆斯·麥德森(James H. Madsen)和威爾斯將角鼻龍下目分為角鼻龍科和雙冠龍科,其中雙冠龍是後者演化支中的唯一物種。[45]

1998年拉曼納和同事指出不只雙冠龍具有頭冠,也曾發現其他有類似頭冠的獸腳類,代表這項特徵並非雙冠龍所獨有,而是作為一個演化支內物種關係的共有特徵。[21]2005年亞當·耶茨(Adam M. Yates)描述了來自南非的龍獵龍,並發現牠與雙冠龍、惡魔龍近緣。他提出的系統發生學分析表示這些物種不屬於腔骨龍超科,而是更衍化的新獸腳類。他指出若雙冠龍比腔骨龍類更為衍化,那一些與後者特徵的相似之處可能是從某些原始獸腳類中遺傳下來的,這代表獸腳類在演化過程的早期階段可能曾經歷過一段「腔骨龍形態」的時期。[46]
2007年內森·迪史密斯(Nathan D. Smith)和同事認為冰脊龍(另一種帶冠獸腳類)與雙冠龍互成姊妹群,並與龍獵龍、中國龍構成一個演化支。該類群較腔骨龍超科衍化、但比角鼻龍類原始,於是使獸腳類的演化進程呈現一種階梯式排列。[47]2012年卡拉諾和同事認為史密斯等人所提出的有冠獸腳類族群僅根據彼此具有類似的頭冠,但骨骼其餘部位缺乏一致的相似性。他們將雙冠龍視為一種腔骨龍科,而冰脊龍和中國龍較進階,屬於堅尾龍類的基礎位置。[13]
2015年罕垂克斯和同事將雙冠龍科定義為包含雙冠龍及龍獵龍兩屬,並提到雖然該演化知的確切位置尚不明確,顯然比腔骨龍超科進階,並與鳥獸腳類互成姊妹群。雙冠龍科與腔骨龍超科的共同特徵包括鼻部下方的凹陷、上頜骨牙齒朝前;而與鳥獸腳類的共有特徵包含上頜骨前面的孔、上頜骨牙齒數量減少。研究認為冰脊龍和中國龍的頭冠都是趨同演化的結果、或是某種從共同祖先遺留下來的特徵。以下演化樹取自該篇研究,數據本身參考更早期的研究:[48]

| 新獸腳類 Neotheropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||
2019年瑪麗安·札訥(Marion Zahner)和威楠·賓克曼(Winand Brinkmann)發現雙冠龍科物種是鳥獸腳類並系群中的姊妹分支,而非一個自然的單系群。但同時仍指出一些分析證實雙冠龍科的有效性,包含雙冠龍、冰脊龍、龍獵龍、可能還有嵌征盗龙作為最基礎的物種;還根據一些下頜特徵提出雙冠龍科的鑑定方法。[49]在2020年馬許和羅威重新描研究中的系統發生學分析,發現雙冠龍的所有標本構成一個單系群,與鳥獸腳類互成姊妹群,並比冰脊龍更衍化。該分析並未支持雙冠龍科的存在,並認為頭冠構造是角鼻龍類和堅尾龍類的祖徵遺傳。[6]
古生物學
食性

威爾斯認為雙冠龍缺乏強勁的咬合力,因鼻下縫導致虛弱。他覺得雙冠龍前上頜骨的牙齒主要用於拉取或撕扯,而非咬合,而後方的上頜骨牙齒用以刺穿和切割。他推測雙冠龍可能是食腐動物而非捕食者,若真能獵殺大型獵物,將是以手腳而非嘴部來進行。威爾斯沒在雙冠龍頭骨中發現任何能使各部位骨頭進行相互移動的證據。[2]1986年羅伯特·巴克卻發現雙冠龍有厚重的頸部與頭骨、及巨大的上排牙齒,適合捕殺大型獵物,且足夠強壯得以攻擊任何已知的早侏羅世植食恐龍。[50]1988年保羅反駁雙冠龍是食腐動物的觀點,並指出完全食腐的陸生動物基本上並不存在。他表示雙冠龍的吻部咬合力比過去認為的要強得多,加上大而細長的上排牙齒相對於指爪來說更為致命。保羅推測牠會捕殺像原始蜥腳形亞目之類的大型獵物,同時又比其他相近尺寸的獸腳類更容易捕捉小型獵物。[38]
2005年法蘭索瓦·特里安(François Therrien)與同事進行了棟樑方程式研究發現雙冠龍下頜的咬合力在咬緊牙齒後快速減低,顯示其下頜骨前部上翹的下巴、玫瑰花座狀的牙齒序列、強化的骨融合區域(類似棘龍科)可能適合用來捕捉及固定相對小型的獵物。雙冠龍下頜骨融合近似於貓科動物及鱷魚制服獵物時透過下頜前部強力咬合的特性。下頜骨所承受的壓力與小型獵物的掙扎一致,可能以切割咬傷的方式來削弱獵物體力直至無力抵抗而以吻部前部捕捉,再來獵物被移動至較大牙齒所在的頜部後方,並以側扁的牙齒來切咬殺死獵物(如同某些鱷魚)。研究人員推測若雙冠龍真以小型獵物為食則其能掠食的體型將受到侷限。[51]
2007年安琪拉·米爾納和詹姆斯·I·科克蘭指出雙冠龍具有食魚的特徵,即頜部末端朝兩側擴張,構成一團互疊牙齒而成的玫瑰花座狀,近似以魚為主食的棘龍科及現代長吻鱷。鼻孔位置也退後至頜骨上,如同鼻孔大幅後退的棘龍科,能防止捕魚過程鼻孔被水濺到;兩者手臂也都很長並有發達的爪子利於捕魚。一座橫跨猶他、亞利桑納、內華達巨大的迪西湖(Lake Dixie),形成於雙冠龍出現之前500至1500萬年,在約消滅地球四分之三物種的三疊紀-侏羅紀滅絕事件後生態圈貧乏的後滅絕時代,為殘存生物提供了豐富的魚類食物來源。[52][15]
2018年馬許和羅威記載了蜥腳形亞目的莎拉龍正模標本骨骼上散佈著疑似的齒痕,可能是由雙冠龍啃食其屍體所留下(巨歿龍太小則不太可能),骨骼位置也可能受到食腐者的移動。其中一例可見於左肩胛骨,其上側表面有個橢圓形凹陷;另外右脛骨前下端也有個大孔洞。挖出莎拉龍正模與副模標本的採集地還包含部分未成年雙冠龍標本。[53]2020年馬許和羅威提出雙冠龍與其他早期獸腳類區別開來的衍化特徵都與體型增加、捕食大型獵物有正相關。儘管兩人仍認同雙冠龍在其棲息環境可能以河流中的魚類及小型獵物為食,他們指出前上頜骨與上頜骨之間的關節是固定的,且比過去所認為得要結實許多,而在捕食大型獵物或食腐的過程中,雙冠龍將以前肢抓取固定。莎拉龍標本上的大型咬痕、脫落的牙齒、同一採集地發現的雙冠龍標本都支持此論點。[6]
2021年馬修·伯朗(Matthew A. Brown)和羅威在一篇文章中表示這些遺骸顯示雙冠龍的下頜強壯得足以刺穿骨頭。從其呼吸系統長入脊椎的肉質氣囊同時對骨骼有著加固與減輕效果,使氣流得以單向流通肺部一如鳥類及鱷魚,將比哺乳類的雙向呼吸還能獲得更多氧氣。單向呼吸顯示代謝率相對較高,等同高度活躍性,代表雙冠龍可能是種快速靈敏的掠食者。布朗和羅威認為雙冠龍是其生態系統的頂級掠食者而非清除者。[15]
活動能力

威爾斯將雙冠龍設想為一個活躍的雙足動物,類似特大號鴕鳥。他認為前肢將作為強有力的武器,強壯而靈活並不用於行走。他指出雙手能夠抓握及揮砍,彼此可互相接觸,並能伸向頸部上方三分之二處。他並推測雙冠龍坐著時,將倚靠在坐骨的大「靴」狀處以及尾巴與腳部。[2]1990年史蒂芬和希薇亞·徹卡司(Stephen and Sylvia Czerkas)兄妹提出雙冠龍纖弱的骨盆用來適應水棲生活,水足以支撐其體重,且還可能是個高效率的游泳者。關於是否生存被侷限在有水環境他們表示存疑,然而後肢比例及力量仍顯示其雙足步行時行動敏捷。[54]保羅假想雙冠龍在向獵物發動攻勢時以尾部進行彈跳,近似袋鼠的行為。[55]
2005年菲爾·森特(Phil Senter)和詹姆斯·羅賓斯(James H. Robins)檢驗了雙冠龍與其他獸腳類前肢的運動範圍。研究顯示雙冠龍能將肱骨向後伸起直至幾乎與肩胛骨平行的角度,但無法前伸到超過垂直角度。肘部能夠接近完全伸展及彎曲,但並非徹底達到。手指似乎無法刻意(向後)過伸(hyperextensible)(超出正常伸屈範圍),但也許能被動屈伸,以抵銷因抓住獵物劇烈掙扎造成的脫臼。[56]2015年森特和羅賓斯的論文針對各屬雙足恐龍進行檢驗,來推測前肢姿勢,其中就包括雙冠龍。肩胛骨維持在接近水平,肘部休止位置接近直角,而手部與下臂間的方向沒有太多偏差。[57]

2018年森特和寇文·蘇黎范(Corwin Sullivan)透過對骨骼的操縱來檢驗雙冠龍前肢關節的運動範圍,以推測各種關於前肢功能的假設。他們還將在檢驗鱷魚屍體觀察到的情況考量進來,其顯示表面包覆軟組織(包括軟骨、韌帶、肌肉)時的肘部運動範圍,遠大於單純只有骨骼的時候。他們還發現雙冠龍的肱骨可以收縮到與肩胛骨近乎平行的位置,並伸直到幾乎完全垂直的角度,而抬高可到達65度角。肘部無法彎曲超過與肱骨夾的角度。手腕的前轉與後轉(Pronation and supination)(尺橈骨以下的手部轉動)被限制住,因尺橈骨關節無法滑動,所以構成手掌朝體內側、彼此平行的狀態。無法前轉的手腕是獸腳類及其他恐龍類群共有的祖徵。手腕運動範圍受限,屈曲時手指分叉並過伸。[58]
森特和蘇黎范得出結論,雙冠龍能用雙手抓握物體;單手抓取或握住小物體;抓住靠近胸下位置的物體;將物體抓到嘴邊;沿胸腔兩側以弧形擺動手臂以進行展示;抓撓胸腹部或另一邊前肢距離最遠的一端;抓住胸部與頸部下方位置的獵物;將物體抱在胸前。雙冠龍無法做出抓鑿、鉤拉、用一隻手兩個指尖夾住物體、將手臂向外伸展以保持平衡、或像現代指猴那般把手或手指伸進小縫中。手指的過伸特性可能防止獵物劇烈掙扎引發脫臼,因為使手指運動程度加大(但對運動沒有實際重要性)。由肩部活動受限、前肢短小來判斷,先接觸獵物的是嘴部而非手部。只有在捕捉小到能擺在胸下位置的獵物時、或透過嘴壓制住更大的獵物時,手部才可能會派上用場。大比例的頭頸部使吻部的延伸範圍遠遠超過手臂可及。[58]

.png.webp)
1971年威爾斯記載的雙冠龍足跡皆位於同一水平面,被描述成像「養雞場大亂步」,很少明確構成移動路線。足跡陷入泥中約5至10公分深,趾痕模糊,其寬度變化代表著泥土沾黏在腳上。印痕也因基質和產生方式不同而有所差異;有時腳直接著地,但經常發生前後滑動。腳趾位置及角度變化也很大,代表勢必相當靈活。雙冠龍足跡中看出第二趾偏轉,基部厚、爪子長直、與趾墊軸線成一直線。其中一個足跡缺少了第二趾爪,可能由受傷所造成。[24]1984年威爾斯將三個個體彼此靠近及附近縱橫交錯足跡的發現解釋為雙冠龍群居活動的證據。[2]給認同小團行動,但指出沒有明確證據來支持,洪水或土石流之類的災難也可能將不同個體的零散骨頭集中成一塊。[12]
2009年米爾納等人檢驗了可能的雙冠龍足跡SGDS 18.T1,包括帶有尾巴拖曳痕跡的典型腳印以及一些異常的靜止痕跡,沉積於湖灘砂岩。足跡始於與岸緣大致平行,然後停在雙腳平行的堤岸前,再來蹲下身體,將蹠骨及坐骨周圍的骨痂碰觸到地表,形成對稱的腳跟與坐骨的圓形印痕。尾巴與身體連接的地方遠離地表,而末端與地表接觸。動物在斜坡上休息,將雙手放到地上靠近腳的位置。休息完後,恐龍拖著腳往前走,腳、蹠骨、坐骨都留下新印痕,手卻沒有。此時右腳踩在原本右手印痕,左腳第二爪由一靜止位到下一靜止位間作了一道拖曳。再過一會,動物左腳在前站起來往前移,完全站直後,牠走過其於暴露的表面,同時尾巴末端留下淺淺的拖痕。[36]
獸腳類中很少發現蹲伏行為。SGDS 18.T1是唯一呈現出清晰手部印痕的獸腳類足跡,提供其如何運用前肢的珍貴資訊。獸腳類的蹲伏被認為與現代鳥類十分相似,顯示早期獸腳類手掌往內側朝向彼此。代表著該姿勢在演化支初期便已出現,並很可能是所有獸腳類共有的特徵。早期的描繪經常將獸腳類手掌朝下,但解剖學研究顯示牠們與鳥類一樣無法將手臂前後轉動。足跡顯示雙腿保持對稱,體重分擔於腳部及蹠骨間,此特徵也可見於平胸鳥類。米爾納等人亦駁斥了偉因斯記載凱恩塔足跡的手掌印痕由四族獸腳類所留下的觀點。偉因斯推測足跡製造者緩慢行走時可用四足步態,手指會習慣性過度伸展,使只有手掌著地。米爾納等人則認為這樣的動作顯得多餘,並推測與SGDS 18.T1類似方式製造足跡但未留下趾痕。[36]
頭冠功能

威爾斯承認關於雙冠龍的頭冠功能都僅止於推測。他認為雖然頭冠中沒有凹槽顯示血管分布,但不排除體溫調節功能的可能性。他還推測頭冠可能用作物種辨識或裝飾物。[2]徹卡司兄妹指出,頭冠無法用於戰鬥,因為結構脆弱容易損傷。他們認為這是調節體溫或吸引配偶的視覺展示物。[54]1990年華特·庫姆斯(Walter P. Coombs)表示頭冠可能具有鮮豔色彩,以增強展示效果。[59]
2011年凱文·帕迪安與約翰·霍納提出許多恐龍的「奇特結構」(如頭冠、褶飾、角、圓顱頂等)主要都用作物種識別,從而反駁其他功能皆無法提供證據支持。他們指出有頭冠的獸腳類標本(包括雙冠龍)太少,無法計算其演化規律、或是否與兩性異形、性成熟有關。[60]同一年羅伯·奈爾(Rob J. Knell)與史考特·山普森在對帕迪安和霍納的回覆中辯論道,不排除物種識別作為恐龍奇特結構的次要功能的可能性,但性擇(展示或爭取配偶的戰鬥)依然是最可能的解釋,有鑑於發育此條件耗費的代價相當高,加上此結構在物種內的變異性似乎也很高。[61]
2013年大衛·伊宏恩(David E. Hone)和戴倫·奈許批判了「物種識別假說」,認為現代沒有物種透過這種結構來進行種間辨識,並且帕迪安和霍納的理論忽略了交互性擇(兩性皆有裝飾物)的可能性。[62]2020年馬許和羅威表示認同雙冠龍的頭冠可能同時在物種識別及性擇中皆有作用,類似某些現代鳥類的例子。[6]對於頭冠內的氣腔是否與前述功能有關,目前尚未明瞭。[15]
發育

威爾斯起初認為雙冠龍較小型的標本是幼體、較大的是成體,後將牠們視為不同物種。[2][7]保羅認為標本間的體格差異可能是兩性異形,腔骨龍似乎有著類似的狀況,同樣體型分為「粗壯」、「纖細」兩種形態,也可能被誤認為不同物種。以此推論較小的雙冠龍標本可能屬於「纖細」形。[38]
2005年泰可斯基根據骨化程度判斷,發現已知的大部分雙冠龍標本是幼年個體,只有最大型的標本為成年。[14]2005年給並未發現保羅提出兩性異形的相關證據(腔骨龍的狀況也僅止於推測),從而將雙冠龍標本間的差異歸因於個體發育過程中及個體間的差異。骨骼並未顯示出兩性異形,但不排除頭冠有的可能性;只是需要更多證據才能支持。[17]耶茨根據幼體標本上的細小鼻冠暫時歸入近緣的龍獵龍,他表示隨動物邁向成年鼻冠將越變越大。[46]
1996年卡赫(J.S. Tkach)發表了雙冠龍組織學研究報告,對UCMP 37303(兩個初始標本中保存較少的那副)的長骨和肋骨進行薄切片。骨骼組織的血管化程度良好,並具類似其他獸腳類與蜥腳形類大椎龍的層狀纖維結構。骨骼織狀結構顯示其生長快速,在雙冠龍的生命初期即可能達到每年增長30至35公斤的速率。[63]
威爾斯發現雙冠龍和其他獸腳類的替生齒發源與骨骼深處,牙齒隨距離齒槽緣越遠尺寸越小。齒槽中通常有二至三顆替生齒,最年輕的牙齒僅有一個小的空心牙冠。替生齒長在舊牙齒外側。當牙齒靠近牙齦線,齒間板的內壁被吸收形成營養凹陷。隨著新牙齒萌發,以齒槽為中心向外移動,營養凹陷閉合。[2]
古病理學

威爾斯在雙冠龍發現許多病徵:正模頸椎神經弓有道溝槽,可能由受傷或擠壓所致;右肱骨有兩個可能由膿瘍或人為製造出的凹陷。威爾斯還稱其左肱骨相較於右側要小且纖弱,但前臂狀況恰好相反。2001年勞弗·莫納提出這是由變動不對稱的異常發育所造成,可能起因於動物族群的壓力,如棲地干擾,可能代表著更強烈的演化壓力。不對稱也可能由動物早期發育的創傷事件所致,其發生機率更為隨機。[2][64]2001年布魯斯·羅斯柴爾德(Bruce Rothschild)和同事對60塊雙冠龍腳部骨頭檢驗是否存在疲勞性骨折(起因於劇烈重複動作),但沒有發現任何跡象。此傷害可能存在於活躍的掠食者。[65]

2016年森特與撒拉·楊思忑(Sara L. Juengst)檢驗了正模病徵,發現其保存了迄今所有已描述獸腳類前肢及胸帶中最多且最多樣的病徵,其中某些尚未在任何其他恐龍發現過。僅知其他有六種獸腳類前肢及胸帶發現過多種病徵。正模有八塊骨頭受損,遠超過其他獸腳類已知最多四塊。左側中,肩胛骨與橈骨骨折,尺骨與外側指骨出現纖維囊腫。右側中,肱骨軸扭曲,橈骨有三個腫瘤,第三掌骨關節面斷裂,第三指第一節變形,該手指永久變形而無法彎曲。肱骨與第三指畸形可能由骨發育不全症造成,過去未曾於非鳥恐龍發現,但已在鳥類發現過。此疾病影響著經歷過營養不良的雛鳥,使其中一邊肢體感到疼痛,從而偏好使用另一邊而導致扭曲。[66]
造成這類創傷的事件次數未知,可能產生於同一次遭遇,如跟別人打架撞到樹或岩石而被爪子刺傷。有鑑於傷口均已痊癒,可以肯定這隻雙冠龍經歷事件後仍存活了很長一段時間,短至數月長至數年。癒合過程必定對前肢捕獵的使用造成限制,因此牠可能承受一段長時間斷食,或靠嘴、腳、或另一邊前肢捕食較小獵物來充飢。據森特和楊思忑所述,恐龍能在多處地點忍受高度疼痛也顯示其為一種高強耐受力的動物。他們並稱恐龍的病徵被低估,雖然威爾斯徹底描述了正模,他只提及發現的其中一項病徵。作者認為這些病徵有時容易被忽略,因為物種描述論文主要關注於特徵而非變異處,或這些異狀難以識別。[66]森特和蘇利文發現正模的病狀改變其右肩與右手第三指的活動範圍,因此對該個體活動範圍的估計可能與健康個體的情況有所落差。[58]
古生態學
地質環境

雙冠龍所知來自凱恩塔層,年代可追溯至早侏羅世的錫內穆期至普林斯巴期,約1億9600萬至1億8300萬年前。[67]凱恩塔層是格蘭峽谷群的一部分,該系列涵蓋了北亞歷桑納、部分東南猶他、西科羅拉多、西北新墨西哥。其主要由兩種岩相構成,一為粉砂岩、另為砂岩沉積物。粉砂岩相在亞利桑那大部分地區發現,而砂岩相則存在於北亞利桑那、南猶他、西科羅拉多、西北新墨西哥。該地層主要為河流沉積,其中粉砂岩相屬於河流系統較緩和、滯慢的部分。凱恩塔沉積層終止於將成為納瓦霍砂岩的風化沙丘區。[68]環境屬於季節性乾旱氣候,沙丘不斷於動物所棲息的潮濕環境中更迭,類似河邊的綠洲,形成一條岸邊長滿針葉樹、並被砂地圍繞的水道。[15]該地層的確切年代尚未知曉,可以判斷的關聯依據包括脊椎動物化石、磁地學、花粉證據等輻射測年方法。[67]根據標本存在於凱恩塔層中的範圍顯示,雙冠龍橫跨了相當久的年代。[6]
凱恩塔層產出一小群但持續增長的生物群。大部分化石來自粉砂岩相。[69]非脊椎動物包含微生物、藻類石灰石、[70]矽化木、[71]植物痕跡、[72]淡水雙殼綱和蝸牛、[68]介形綱[73]以及無脊椎動物生痕化石。[70]其餘已知大多數生物都是脊椎動物,包含遺骸與生痕。有遺骸的脊椎動物包含[69]弓鯊科、未定硬骨魚、肺魚、[71]蠑螈、[74]前躍蛙、蚓類的始蚓、凱恩塔龜、一種喙頭目、蜥蜴、[75]數種早期鱷形超目如卡索亞鱷、始氣腔鱷、凱恩塔鱷、原鱷、翼龍類的喙頸龍。除了雙冠龍之外已知還有數種恐龍:獸腳類巨歿龍、[14]凱恩塔獵龍;[76]蜥腳形亞目莎拉龍;[77]一種畸齒龍科;裝甲類小盾龍。合弓綱有三瘤齒獸科丁尼齒獸、凱恩塔獸、小駝獸、摩根錐齒獸類;可能還有早期的真正哺乳類丁尼獸與一種賊獸目。大多發現於亞利桑那的金泉附近。[69]脊椎動物生痕化石有糞化石;獸孔目、類似蜥蜴的動物、數種恐龍足跡。[70][78]
埋藏學
威爾斯大略描述了初始標本的埋藏方式(即屍體分解與化石化過程的變化)。正模標本發現時倒向右側,頸部與尾部後彎,呈現出恐龍遺骸常見的「死亡姿勢」。此姿勢現認為由角弓反張(屍體痙攣)造成,也可能是由屍體嵌入沉積物所致。背部挺直,而最後的背椎朝左側偏轉。尾椎由骨盆不規則延展開來,腿部則關節相連幾乎無位移。威爾斯提出結論為標本於死亡原位保存,未經移動,其中正模似乎曾受啃食者破壞,其頭骨及脊椎被壓碎。[2][79]給指出其2001年描述的標本顯示出被河流搬運的跡象,因沒有一個是完整的,可能經過搬移了一段距離,或搬移前已躺在地表一段時間而被風化。由發現的各種碎片及斷裂跡象判斷可能由洪水搬運。[12]
文化意義
影視娛樂

雙冠龍出現在作家麥可·克萊頓1990年的小說侏羅紀公園與其1993年由史蒂芬·史匹柏執導的改編電影。侏羅紀公園系列的雙冠龍在其幕後花絮書籍中被提到是「唯一嚴重偏離科學準確性的動物」,以及在由史丹·溫斯頓工作室一本關於製作電腦特效的書中提到是電影系列的所有恐龍中「虛構程度最高的」。小說中克萊頓賦予恐龍噴吐毒液的能力,以彌補下頜纖弱的弱點。電影藝術家則還添加了頸部的褶皺皮膜,攻擊時會如同現代傘蜥蜴將皮膜展開震動。為避免與片中迅猛龍混淆,雙冠龍的體型被縮減至只有1.2公尺高,而非現實中古生物學家估計的3公尺。電影的雙冠龍被暱稱為「噴毒者」(the spitter)。片中場景是以木偶戲製作而成,需要一個完整的身體及三個可互換的頭來做出劇本所要求的動作。而也製作了個別的腿供跳躍畫面使用。與電影中多數其他恐龍最不同的是,雙冠龍登場時沒有使用到任何電腦圖像。[80][81][82]

1996年地質學家班寧頓(J. Bret Bennington)指出雖然雙冠龍可能沒有頸褶也不會吐毒液,但咬可能帶有毒性,一如科摩多龍。他認為增加毒液的想法就像同樣未知的皮膚顏色假想一樣是可行的。若恐龍帶有頸褶,骨骼應會發現相關證據,如以堅固結構來支撐,或在附著處有用來移動的肌肉痕跡。他還補充道若頸褶確實存在則功能並非來嚇阻獵物,而是競爭者;他推測電影情節的角色可能是戴上連帽而讓雙冠龍認為是威脅而作出反應。[83]1997年一本討論侏儸紀公園科學的書評中,彼得·多德森同樣指出電影中雙冠龍的錯誤比例與毒液、頸褶的可能性。[84]2014年巴克指出電影的雙冠龍缺乏明顯的上頜凹槽,總結道電影製作人在創造可怕嵌合體分面做得很好,但警告不能拿來當作真正動物的教材。[85]布朗和馬許表示雖然這些特徵是虛構,但參考真實動物生理學而增加了可信度。[15]威爾斯本人對雙冠龍登場於侏儸紀公園表示興奮,就算發現其中存在小問題和不準確性,依然欣賞這部電影,也很高興「恐龍成為國際知名演員」。[86]
現實象徵
_-_close-up.JPG.webp)
_-_prints.JPG.webp)
根據納瓦霍神話,屠殺的怪物屍體被打入土中,但無法消滅,化石便成為傳統上對其遺骸的解釋。自19世紀以來納瓦霍人曾協助古生物學家定位化石,傳統觀念認為怪物魂魄依然殘留於部分掩埋的屍體中,且必須透過強力儀式將其鎮壓在原地。同樣地,有些民眾認為他們親戚的骨骸會隨恐龍化石一同被挖出,而將之移除會造成對這些生命生前的不尊重。[87]2005年歷史學家亞淳·梅約(Adrienne Mayor)表示威爾斯曾提到最早挖掘雙冠龍的過程中,納瓦霍人威廉斯幾天後就消失無蹤,並推測可能是因為他覺得挖掘工作用細刷仔細清修「有失其尊嚴」。梅約卻指出納瓦霍人實際上熱衷於珠寶及繪畫等需要細心的工作,而威廉斯的退出也許只是傳統上對骷髏出現所引起的焦慮。梅約亦提到1940年代曾發生過一名納瓦霍人協助五角龍骨骼的挖掘,前提是他不用碰觸到骨骼本身,並於只剩少許塵土覆蓋骨骼時便離開現場。[87]在1994年的書裡,威爾斯提到威廉斯在幾天後曾帶著兩個納瓦霍女性回歸,並稱「那不是男人的工作,是Squaw(婦女的貶稱)的工作」。[9]
出產雙冠龍化石的亞歷桑納懸崖,亦可見到其上覆蓋著古普韋布洛人的岩刻,當地縱橫交錯的足跡被納瓦霍人稱為「Naasho’illbahitsho Biikee」,意即「大蜥蜴足跡」。據梅約稱,納瓦霍人常舉行儀式向這些怪物足跡獻祭。霍皮族和祖尼人服裝和石飾藝術上的三趾腳印,靈感也可能來自雙冠龍之類的足跡。[87]2017年雙冠龍被指定為康乃狄克州的州恐龍,隨後2019年成為新的州官方預算的一部分。之所以選擇雙冠龍是因為1966年在岩丘91號州際公路建設期間發現了認為是類似動物的足跡,該六道足跡於1991年歸入巨型真雷龍足跡屬,其發現地原是一座三疊紀的湖泊,就在確認了當地的重要意義後,高速公路因而改道,並在原地設立了一座州立公園名為恐龍州公園。1981年一座雙冠龍雕像被捐贈給公園,成為此物種首個真實比例復原模型。[38][88][89]
1998年一名9歲男童提名雙冠龍作為亞歷桑納州的州恐龍。但立法者提議使用索諾拉龍,宣稱雙冠龍並非亞歷桑納州特有。有人曾提出可同時承認兩種恐龍的折衷方案,但後來撤銷了,因為意識到雙冠龍化石未經納瓦霍保留地許可被帶走,且本身早已不位在亞歷桑納州。2018年一名11歲男童再次提議索諾拉龍應作為亞歷桑納州恐龍。納瓦霍族官員隨即展開討論如何讓化石歸還回來。[90][91]據梅約所述,一名納瓦霍人表示不再堅持取回化石,但想了解為何沒有製作模型以留下骨頭本身,因為能留在地下最好,還可以建造博物館讓民眾可以到原址參觀。[87]在納瓦霍保留地礦產部的許可下,納瓦霍保留地的雙冠龍田野考察工作進一步展開。[15]
參考來源
- . Oxford Dictionaries. Oxford University Press.
- Welles, S.P. . Palaeontographica Abteilung A. 1984, 185: 85–180.
- Welles, S.P. . Bulletin of the Geological Society of America. 1954, 65 (6): 591–598. Bibcode:1954GSAB...65..591W. doi:10.1130/0016-7606(1954)65[591:NJDFTK]2.0.CO;2.
- Welles, S.P.; Guralnick, R.P. . ucmp.berkeley.edu. University of California, Berkeley. 1994 [2018-02-13]. (原始内容存档于2017-11-08).
- Naish, D. . London, UK: A & C Black Publishers Ltd. 2009: 94–95. ISBN 978-1-4081-1906-8.
- Marsh, A.D.; Rowe, T.B. . Journal of Paleontology. 2020, 94 (S78): 1–103. S2CID 220601744. doi:10.1017/jpa.2020.14
 .
. - Welles, S.P. . Journal of Paleontology. 1970, 44 (5): 989. JSTOR 1302738.
- Welles, S.P.; Guralnick, R.P. . ucmp.berkeley.edu. University of California, Berkeley. 1994 [2018-02-13]. (原始内容存档于2017-08-02).
- Psihoyos, L.; Knoebber, J. . London, UK: Cassell. 1994: 86–89. ISBN 978-0679431244.
- Glut, D.F. . Jefferson: McFarland & Company, Inc. 1997: 347–350. ISBN 978-0786472222.
- Rauhut, O.W. . Special Papers in Palaeontology. 2004, 69: 213 [2022-01-30]. (原始内容存档于2022-03-24).
- Gay, R. . Western Association of Vertebrate Paleontologists Annual Meeting 1. Mesa, Arizona: 1. 2001.
- Carrano, M.T.; Benson, R.B.J.; Sampson, S.D. . Journal of Systematic Palaeontology. 2012, 10 (2): 211–300. S2CID 85354215. doi:10.1080/14772019.2011.630927.
- Tykoski, R.S. (学位论文). University of Texas: 1–232. 2005 [2022-01-31]. (原始内容存档于2020-07-29) –UT Libraries: Electronic Theses and Dissertations.
- Brown, M. A.; Marsh, A. D. . Scientific American. 2021, 324 (1): 46–53 [2022-01-30]. doi:10.1038/scientificamerican0121-46 (不活跃 October 31, 2021). (原始内容存档于2021-01-08).
- Pickrell, J. . National Geographic. Science. 2020-07-07 [2020-07-12]. (原始内容存档于2021-02-17).
- Gay, R. . Carpenter, K. (编). . Bloomington, IN: Indiana University Press. 2005: 277–283. ISBN 978-0-253-34539-4.
- Mortimer, M. . Theropoddatabase.blogspot.com. May 2010 [2017-12-29]. (原始内容存档于2017-12-29).
- Hendrickx, C.; Mateus, O.; Evans, Alistair Robert. . PLOS ONE. 2014, 9 (3): e88905. Bibcode:2014PLoSO...988905H. PMC 3943790 . PMID 24598585. doi:10.1371/journal.pone.0088905 .
- Hu, S. . Vertebrata PalAsiatica. 1 1. 1993, 31: 65–69 (中文).
- Lamanna, M.C.; Holtz, T.R., Jr.; Dodson, P. . Journal of Vertebrate Paleontology. Abstracts of Papers, Fifty-Eighth Annual Meeting, Society of Vertebrate Paleontology. 1998, 18 (3): 57–58. JSTOR 4523942.
- Xing, L.; Bell, P.R.; Rothschild, B.M.; Ran, H.; Zhang, J.; Dong, Z.; Zhang, W.; Currie, P.J. . Chinese Science Bulletin. 2013, 58 (16): 1931. Bibcode:2013ChSBu..58.1931X. doi:10.1007/s11434-013-5765-7 .
- Wang, Guo-Fu; You, Hai-Lu; Pan, Shi-Gang; Wang, Tao. . Vertebrata PalAsiatica. 2017, 55 (2): 177–186 [2022-01-30]. (原始内容存档于2021-08-19).
- Welles, S.P. . Plateau. 1971, 44: 27–38.
- Morales, M.; Bulkley, S. . Museum of Northern Arizona Bulletin. 1996, 60: 143–145.
- Gierliński, G. . Palaeogeography, Palaeoclimatology, Palaeoecology. 1991, 85 (1–2): 137–148. Bibcode:1991PPP....85..137G. doi:10.1016/0031-0182(91)90030-U.
- Gierliński, G.; Ahlberg, A. . Ichnos. 1994, 3 (2): 99. doi:10.1080/10420949409386377.
- Gierliński, G. . Museum of Northern Arizona Bulletin. 1996, 60: 179–184.
- Kundrát, M. . Journal of Experimental Zoology Part B: Molecular and Developmental Evolution. 2004, 302B (4): 355–364. PMID 15287100. doi:10.1002/jez.b.20014.
- Lockley, M.; Matsukawa, M.; Jianjun, L. . Ichnos. 2003, 10 (2–4): 169–177. S2CID 128759174. doi:10.1080/10420940390256249.
- Martin, A.J.; Rainforth, E.C. . Geological Society of America. Abstracts with Programs. 2004, 36 (2): 96. (原始内容存档于2004-05-31).
- Weems, R.E. . le Tourneau, P.M.; Olsen, P.E. (编). 2. New York: Columbia University Press. 2003: 293–313. ISBN 978-0231126762.
- Rainforth, E.C. . Palaeontology. 2003, 46 (4): 803–838. doi:10.1111/1475-4983.00320 .
- Weems, R.E. . New Mexico Museum of Natural History and Science Bulletin. 2006, 37: 369–378.
- Lucas, S.G.; Klein, H.; Lockley, M.G.; Spielmann, J.A.; Gierlinski, G.D.; Hunt, A. P.; Tanner, L.H. . New Mexico Museum of Natural History and Science Bulletin. 2006, 37 [2022-02-13]. 265. (原始内容存档于2016-10-04).
- Milner, Andrew R.C.; Harris, J.D.; Lockley, M.G.; Kirkland, J.I.; Matthews, N.A.; Harpending, H. . PLOS ONE. 2009, 4 (3): e4591. Bibcode:2009PLoSO...4.4591M. PMC 2645690 . PMID 19259260. doi:10.1371/journal.pone.0004591 .
- Weems, R.E. . Ichnos. 2019, 26 (3): 187–215. S2CID 133770251. doi:10.1080/10420940.2018.1532902.
- Paul, G.S. . New York, NY: Simon & Schuster. 1988: 258, 267–271. ISBN 978-0-671-61946-6.
- Holtz, T.R., Jr. . New York, NY: Random House. 2012: 81. ISBN 978-0-375-82419-7.
- Paul, G.S. . Princeton, NJ: Princeton University Press. 2010: 75. ISBN 978-0-691-13720-9.
- Tykoski, R.S.; Rowe, T. . Weishampel, D.B.; Dodson, P.; Osmolska, H. (编). 2. Berkeley, CA: University of California Press. 2004: 47–70. ISBN 978-0-520-24209-8.
- Welles, S.P. . Journal of Paleontology. 1983, 57 (2): 401. JSTOR 1304663.
- Welles, S.P.; Long, R.A. [Annals of the South African Museum]. Annale van die Suid-Afrikaanse Museum. 1974, 64: 191–218 [2022-02-08]. ISSN 0303-2515. (原始内容存档于2022-02-08).
- Holtz, T.R., Jr. . Journal of Paleontology. 1994, 68 (5): 1100–1117. JSTOR 1306180. doi:10.1017/S0022336000026706.
- Madsen, J.H.; Welles, S.P. . Utah Geological Survey. 2000: 1–89 [2022-02-08]. 41293. (原始内容存档于2018-01-13).
- Yates, A.M. . Palaeontologia Africana. 2005, 41: 105–122 [2022-02-08]. ISSN 0078-8554. (原始内容存档于2022-02-08).
- Smith, N.D.; Makovicky, P.J.; Hammer, W.R.; Currie, P.J. . Zoological Journal of the Linnean Society. 2007, 151 (2): 377–421. doi:10.1111/j.1096-3642.2007.00325.x .
- Hendrickx, C.; Hartman, S.A.; Mateus, O. . PalArch's Journal of Vertebrate Palaeontology. 2015, 12 (1): 73 [2018-01-13]. (原始内容存档于2018-06-22).
- Zahner, M.; Brinkmann, W. . Nature Ecology & Evolution. 2019, 3 (8): 1146–1152. PMC 6669044 . PMID 31285577. doi:10.1038/s41559-019-0941-z.
- Bakker, R.T. . New York, NY: William Morrow. 1986: 263–264. ISBN 978-0-8217-5608-9.
- Therrien, F.; Henderson, D.; Ruff, C. . Carpenter, K. (编). . Indiana University Press. 2005: 179–230. ISBN 978-0-253-34539-4.
- Milner, A.; Kirkland, J. (PDF). Survey Notes of the Utah Geological Survey. 2007, 39: 1–3 [2022-03-23]. (原始内容 (PDF)存档于2017-11-04).
- Marsh, A.D.; Rowe, T.B. . PLOS ONE. 2018, 13 (10): e0204007. Bibcode:2018PLoSO..1304007M. PMC 6179219 . PMID 30304035. doi:10.1371/journal.pone.0204007 .
- Czerkas, S.J.; Czerkas, S.A. . Limpsfield: Dragons’ World. 1990: 208. ISBN 978-0-7924-5606-3.
- Paul, G.S. (编). . . New York, NY: St. Martin's Press. 2000: 216. ISBN 978-0-312-31008-0.
- Senter, P.; Robins, J.H. . Journal of Zoology. 2005, 266 (3): 307–318. doi:10.1017/S0952836905006989.
- Senter, P.; Robins, J.H. . PLOS ONE. 2015, 10 (12): e0144036. Bibcode:2015PLoSO..1044036S. PMC 4684415 . PMID 26675035. doi:10.1371/journal.pone.0144036 .
- Senter, P.; Sullivan, C. . Palaeontologia Electronica. 2019. doi:10.26879/900 .
- Coombs, W.P. . Weishampel, D.B.; Osmolska, H.; Dodson, P. (编). 1st. Berkeley, CA: University of California Press. 1990: 42. ISBN 978-0-520-06727-1.
- Padian, K.; Horner, J.R. . Journal of Zoology. 2011, 283 (1): 3–17. doi:10.1111/j.1469-7998.2010.00719.x .
- Knell, R.J.; Sampson, S. (PDF). Journal of Zoology. 2011, 283 (1): 18–22 [2022-02-21]. doi:10.1111/j.1469-7998.2010.00758.x. (原始内容存档 (PDF)于2018-07-22).
- Hone, D.W.E.; Naish, D. . Journal of Zoology. 2013, 290 (3): 172–180. doi:10.1111/jzo.12035 .
- Tkach, J.S. . . Fifty-sixth Annual Meeting, Society of Vertebrate Paleontology. Abstracts of Papers 16 (3). American Museum of Natural History, New York, New York: 1A–80A. 1996-10-19. JSTOR 4523755.
- Molnar, R.E. . Tanke, D.H.; Carpenter, K. (编). . Bloomington, IN: Indiana University Press. 2001: 337–363. ISBN 978-0-253-33907-2.
- Rothschild, B.; Tanke, D.H.; Ford, T.L. . Tanke, D.H.; Carpenter, K. (编). . Bloomington, IN: Indiana University Press. 2001: 331–336. ISBN 978-0253339072.
- Senter, P.; Juengst, S.L.; Heymann, D. . PLOS ONE. 2016, 11 (2): e0149140. Bibcode:2016PLoSO..1149140S. PMC 4765892 . PMID 26909701. doi:10.1371/journal.pone.0149140 .
- J. M., Clark; Fastovsky, D.E. . Fraser, N.C.; Sues, H.-D. (编). . Cambridge, UK: Cambridge University Press. 1986: 285–301. ISBN 978-0-521-36779-0.
- Harshbarger, J. W.; Repenning, C. A.; Irwin, J. H. . New Mexico Geological Society. 1957, 291: 98–114 [2022-03-31]. (原始内容存档于2020-02-16).
- Lucas, S.G.; Heckert, A. B.; Tanner, L.H. . New Mexico Museum of Natural History Bulletins. 2005, 29: 95–104 [2022-03-31]. (原始内容存档于2016-11-18).
- Luttrell, P.R.; Morales, M. . Museum of Northern Arizona Bulletin. 1993, 59: 111–127.
- Jenkins, F.A.; Shubin, N. H. . Journal of Vertebrate Paleontology. 1998, 18 (3): 495–510. doi:10.1080/02724634.1998.10011077.
- Rigby, J.K.; Hamblin, W.K.; Matheny, R.; Welsh, S.L. (PDF). Brigham Young University Research Studies, Geology Series. 1971, 18 (2): 7 [2022-03-31]. (原始内容 (PDF)存档于2020-11-25).
- Lucas, S.G.; Tanner, L.H. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2007, 244 (1–4): 242–256. Bibcode:2007PPP...244..242L. doi:10.1016/j.palaeo.2006.06.030.
- Curtis, K.; Padian, K. . PaleoBios. 1999, 19 (2): 19–37.
- Jenkins, F. A.; Crompton, A. W.; Downs, W. R. . Science. 1983, 222 (4629): 1233–1235. Bibcode:1983Sci...222.1233J. PMID 17806725. S2CID 35607107. doi:10.1126/science.222.4629.1233.
- Gay, R. . Gay, R. (编). . Lulu Press. 2010: 27–43. ISBN 978-0557466160.
- Rowe, T.B.; Sues, H.-D.; Reisz, R.R. . Proceedings of the Royal Society B: Biological Sciences. 2010, 278 (1708): 1044–1053. PMC 3049036 . PMID 20926438. doi:10.1098/rspb.2010.1867.
- Hamblin, A.H.; Foster, J.R. . Sprinkel, D.A.; Chidsey, T.C., Jr.; Anderson, P.B. (编). . Salt Lake City: Utah Geological Association. 2000: 557–568. ISBN 978-1882054107.
- Reisdorf, A.G.; Wuttke, M. . Palaeobiodiversity and Palaeoenvironments. 2012, 92 (1): 119–168. doi:10.1007/s12549-011-0068-y.
- Crichton, M. . London: Random Century Group. 1990: 76–78. ISBN 978-0-394-58816-2.
- Shay, D.; Duncan, J. . New York: Boxtree Ltd. 1993: 24, 35–36, 113. ISBN 978-1-85283-774-7.
- Duncan, J. . London, UK: Titan Books. 2007: 177–178. ISBN 978-1-84576-150-9.
- Bennington, J.B. . American Paleontologist. 1996, 4 (2): 4–7 [2017-12-28]. (原始内容存档于2018-10-02).
- Dodson, P. . Science. 1997, 277 (5326): 644–645. JSTOR 2893254. S2CID 178195275. doi:10.1126/science.277.5326.644b.
- Bakker, R.T. . blog.hmns.org. The Houston Museum of Natural Science. 2014 [2020-08-18]. (原始内容存档于2020-09-21).
- Welles, S.P.; Guralnick, R.P. . ucmp.berkeley.edu. University of California, Berkeley. 1994 [2018-02-13]. (原始内容存档于2018-01-04).
- Mayor, A. . . Princeton, NJ: Princeton University Press. 2005: 130–143. ISBN 978-0691130491. JSTOR j.ctt4cgcs9.10.
- Stone, M. . CTBoom.com. 2017 [2018-02-13]. (原始内容存档于2017-09-25).
- Rogers, O. . Connecticut History. 2016 [2018-01-03]. (原始内容存档于2022-04-11).
- Moeser, C. . The Arizona Republic. Deseret News. 1998-04-26 [2020-05-12]. (原始内容存档于2022-05-29).
- Gebers, S. . The Republic. 2018-02-19 [2018-02-20] –AZ Central.