蝴蝶
蝴蝶是一類常於日間飛行的昆蟲,生物分類學中屬於鱗翅目中名为鳳蝶總科(學名:Papilionoidea,又名真蝶總科)的一個總科級演化支[2]。其下共有7個科,包含了大約18,800種 [3]。如同其他昆蟲,蝴蝶也屬於節肢動物,體表具有分節的外骨骼,成蟲身體也分為頭、胸、腹三個部份,胸部長有兩對翅膀,翅膀上各式各樣的色彩和斑紋是由翅膀上的鱗片組成。蝴蝶是完全變態昆蟲,其一生包括卵、幼蟲、蛹與成蟲四個階段。世上體型最大的蝴蝶為亞歷山大鳥翼鳳蝶,展翅寬達280毫米[4][5],體型最小的蝴蝶是褐小灰蝶,只有16毫米[6]。
| 蝴蝶 | |
|---|---|
 | |
| 玉帶鳳蝶 Papilio polytes | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 节肢动物门 Arthropoda |
| 纲: | 昆虫纲 Insecta |
| 目: | 鱗翅目 Lepidoptera |
| 亚目: | 有喙亞目 Glossata |
| 演化支: | Coelolepida |
| 演化支: | 肌旋喙類 Myoglossata |
| 演化支: | 新鱗翅類 Neolepidoptera |
| 下目: | 韁翅下目 Heteroneura |
| 演化支: | 真鱗翅類 Eulepidoptera |
| 演化支: | 真異脈類 Euheteroneura |
| 演化支: | 雙孔類 Ditrysia |
| 演化支: | 短突雙孔類 Apoditrysia |
| 演化支: | 螟蛾類 Obtectomera |
| 总科: | 鳳蝶總科 Papilionoidea |
| 科 | |
| 多样性 | |
| 共有 7 個科 約 18,800 種 | |
分佈
蝴蝶廣泛分佈世界各地的陸上地方,除了南極洲以外皆有分佈,熱帶地區物種多樣性最高,溫帶及寒帶地區也有許多種類棲息。世上共有約18,500種蝴蝶,新北界有約775種,新熱帶界約7700種,古北界約1575種,埃塞俄比亞界約3650種,東洋界及澳新界共約4800種[7]。在中國大陆記錄了約2100種;台灣約370種;香港約260種。
- 生境
蝴蝶的生境十分廣泛,但主要都是圍繞牠們賴以為生的植物,包括蜜源植物和寄主植物。前者是蝴蝶成蟲攝取花蜜的開花植物,後者是幼蟲食用的植物。
生命史
- 可參閱:完全變態
蝴蝶是完全變態的昆蟲,牠們的一生有卵、幼蟲、蛹和成蟲四個階段。這種生存方式有更強的競爭性,例如幼蟲和成蟲間完全不同的食性能減少種內競爭、一生特化成各個階段能更有效率地完成不同階段的任務:幼蟲有發達的消化系統使能不斷進食並貯存能量、蛹期不用進食能渡過不利生長的冬季、成蟲長有翅膀和成熟的生殖系統能有效找尋配偶並繁殖後代。
蝴蝶壽命一般有1至4個月,不過不同季節和種類其壽命都不同。在溫暖和食物充足的時候蝴蝶能快速成長,壽命會較短,反之亦然。一般成蝶只有兩個月壽命,但以成蟲越冬的斑蝶,成蟲壽命可以長達數個月。
卵
_laying_eggs_I_IMG_9965.jpg.webp)
產卵方式依種類不同,產卵位置可以是在寄主植物葉片、嫩芽、枝條,休眠芽附近,或是在寄主植物附近的其他植物體或地面上。也有將卵產在樹皮裂缝,花苞及新芽孔隙的情形。卵粒單產或數粒成一群,也有一次產數百粒成卵塊的種類。通常卵粒藉由雌蝶副腺分泌物黏著在物體上,也有完全隱藏在泡狀或膠狀物中的情形,更有少數種類會有直接將卵粒拋置地面的特殊習性。
幼蟲
卵剛孵化成蟲時稱作一齡蟲,幼蟲一般從卵頂部嚙破卵殼出來,但是卵粒產在裂縫與孔隙的種類有些具有幼蟲從側面破卵而出的適應性,甚至有針對卵蜂演化出加厚卵殼而幼蟲從卵穿過葉片脫出的種類。有部分種類在孵化後幼蟲會啃食卵殼,使不會浪費資源,又可以防止空卵殻暴露了自己的位置,吃過卵殼後會不斷吃寄主植物。極少數蝴蝶的幼蟲會吃其他生物,例如灰蝶中雲灰蝶亞科的蝶種都是肉食性,如蚜灰蝶的幼蟲以蚜蟲為食、熙灰蝶幼蟲以介殼蟲為食[8]。直至下一個階段脫皮轉齡,幼蟲會停止活動及進食,作脫皮轉齡的預備。脫皮開始時,幼蟲身體伸縮,將舊皮從頭部脫到尾部,最後把頭部的外殻脫下便完成脫皮。幼蟲每次脫皮為一齡,會脫皮4至6次。

老熟幼蟲會停止進食,到處爬行尋找結蛹的地方,通常會選擇停留在葉底。為了確保羽化成蝴蝶時能順利展翅,幼蟲會拉長身子量度空間是否足夠。決定好結蛹地點後,幼蟲會先吐絲作墊,用尾足鉤著絲墊固定位置,然後不斷地向左右吐絲,造成一條粗帶固定蟲體,形成前蛹。大約一天後便開始化蛹,這時,前蛹會有間歇性的伸縮及膨脹活動。隨後伸縮及膨脹的次數會漸趨頻密,表皮從胸前裂開,並迅速被推至腹部末端。最後,末端臀棘鉤著絲墊,將最後一張皮也脱下時便完成化蛹。
蛹
蛹的表面看似靜止不動,其實牠第四至第六腹節是能夠活動的,在這階段蛹內會進行複雜的重組過程,把幼蟲時的器官分解,並組成成蟲的身體。約十天後,成蟲的器官會在蛹內發育完成,接著便是羽化。
成蟲
蝴蝶的羽化一般在深夜至清晨進行。成蟲開始羽化時,蛹殼頭部和胸背呈十字形裂開,頭、胸及背部一一露出,接著前足伸出,並將皺摺的翅膀及鼓脹的腹部拉出蛹殼外。剛爬出的成蟲通常會附在蛹殼下,或在其他穩固位置將身體懸掛,引導腹部的血淋巴體液注入翅脈,流進翅膀,並將摺皺的翅膀迅速撐開。當雙翅伸展後,多餘的體液會由肛門排出。假如此時蝴蝶掉在地上或有障礙物令翅膀不能順利伸展,牠便會永久失去飛行能力。待翅膀變硬後,成蟲便具備飛行的能力。
_mating%252C_male_(l)_and_female_(r).jpg.webp)
羽化後的成蟲,無論在外形和結構都與幼蟲全不相同,也是我們最熟悉的蝴蝶。成蟲所有器官已發育成熟,具備交配與產卵的能力,繁殖下一代,通常蝴蝶一生只會交配一次。雄蝶會找尋雌蝶交配,交配後的雌蝶會找尋寄主植物產卵,由下一代延續蝴蝶的生命史[9]。
形態
頭部
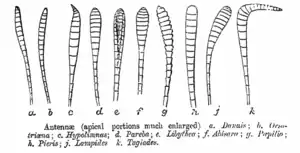

蝴蝶頭部呈圓球形或平球形,表面密被鱗毛,左右兩側各具一發達的複眼,由上萬個六角形小眼組成,每個小眼能看到所見的對象的一個小光點,無數小眼所見的無數小光點造成鑲嵌而成的影像。許多種類在小眼間生有細毛,複眼是否具毛以及其被毛狀態在分類上頗有價值。頭部背側和複眼之間有一對细長,生許多有感覺器官的觸角,分為柄節、梗節和鞭節。鞭節在外觀上分為許多小節,但是這些小節內部沒有獨立運作的肌肉連繋。鞭節末端的膨大部分可稱為錘部,錘部成圓柱狀或壓扁狀,錘部末端稱為尖頂,在弄蝶科格外發達。觸角表面被覆鱗片,但在末端有裸露部分,又稱為裸節。在觸角後方有毛叢,稱為毛隆。口器位於頭部下側,高度特化以利吸食流體食物,依種類與性別不同會吸食花蜜、腐果、動物排遺、溪流邊與地面水分等。口器基本上只餘下唇鬚及小顎外葉,上唇和上領部分則已退化消失。下唇鬚呈片狀,能前伸或上舉,在喙蝶亞科特別明顯。小顎外葉形成兩根半管,左右相互嵌合成一形如彈簧的吸管,休息時螺旋狀捲縮在頭的下面,應用時又可伸直,以虹吸力吸花蜜等汁液。
胸部
蝴蝶胸部由三節組成,分別為前胸、中胸與後胸。前胸窄小,兩側生有氣孔,背側有骨化或膜質構造,稱為背翼。胸部具有兩對翅,位於中胸者為前翅,後胸者為後翅。前翅基部有用以保護翅基關節之骨片,稱為肩板。胸部各節亦分別有一對足,由前至後分別為前足、中足及後足[10]。足主要分為基節、轉節、腿節、脛節及跗節五節,其上覆有鱗片與刺毛,脛節上常具成列小刺。中足及後足脛節上常具一或兩對具關節的距。鳳蝶與弄蝶前足脛節上具有稱為前脛突的特殊構造,生有梳狀刺毛[11]。跗節通常分為五節小跗節,末端生有一對彎曲的爪,但在蛺蝶科及灰蝶科雄蝶跗節常癒合成圓筒狀,不分節,末端無爪的構造。
腹部
蝴蝶腹部包含消化、排泄及繁殖器官,由10節腹節組成,各腹節由骨化程度較高的一片背板及腹板,以及兩側兩片骨化程度弱的側板所構成,第1至7腹節除了第1腹節缺乏腹板以外,其餘各節形態簡單,缺乏特化構造,但在側板上具有氣孔。雌蝶第8至10腹節以及雄蝶第9至10腹節高度特化,形成外生殖器官,這些構造與部分雌蝶部分内生殖器構造合稱為交尾器。
翅膀

蝴蝶翅膀構造獨特,前翅翅形常接近三角形,後翅則接近扇形,兩者翅前方邊緣稱為前緣,後方邊緣稱為後緣,與身體相接處稱為翅基,而遠離身體的邊緣稱為外缘。前緣與外緣相接處稱為翅頂,後緣與外緣相接處稱為臀角。
翅膀呈膜質,有骨化之翅脈,分別有六組縱脈,由前至後分別為前緣脈(C)、亞前緣脈(Sc)、脛脈(R)、中脈(M)、肘脈(Cu)及臀脈(A)。蝴蝶的前緣脈明顯退化,僅在活體前翅前緣有模糊的痕跡,在乾燥標本上則完全消失。肘脈可分為前肘脈(CuA)與後肘脈(CuP),不過在蝴蝶中僅有鳳蝶科仍具有後肘脈翅脈間的區域稱為翅室。翅室名稱跟隨其前方翅脈名稱,如M1脈後方的翅室即為M1室。在脛脈與肘脈間形成一空室,稱為中室,中室外端有時具一橫脈稱為中室端脈,中室端脈存在時稱為中室封閉,而中室端脈缺少則稱為中室開放。此外,後翅Sc+R1脈近基部位置常有一稱為肩脈(hm)的游離短脈。

蝴蝶翅面上布滿鱗片,又稱作鱗粉,呈覆瓦狀排列。鱗片基部有一小柄,藉以附著於翅膜上細微窩孔上。鱗片壁及孔隙中包含由幼蟲時期從寄主植物獲得的成分,成蝶自行代謝出的產物,或是代謝廢物等來源之色素,這類色彩稱為「化學色」或「色素色」,部分蝶種的鱗片上具有各種物理甚至近乎熒光特化結構,可造成光線干涉等物理效應形成金屬光澤的效果,這類色彩稱為「物理色」或「結構色」[12]。

許多雄蝶及少數雌蝶在翅膀上長有用作釋放與求偶有關的信息素的特化鱗,稱為香鱗。香鱗有的散布在翅面上,有的聚成一片形成「性標」,有的則位於特化的袋狀構造或翅褶內。
- 多型性
當同一種蝴蝶出現兩種或以上的型態,便稱為多型性。與個體差異不同,多型性的型態差異較大,各型態亦恆常發現。多型性包括兩性異形、季節異形[13]等型態。兩性體亦會罕有地發生,即同一隻蝴蝶擁有兩性的性器官,左右翅膀的花紋都不一樣,甚至形狀不同。

.jpg.webp)

幼期
蝴蝶的卵具多變化的形態,外部具有卵殼,卵殼頂部中央有稱為精孔的小孔。
蝴蝶的幼蟲身體常呈圓筒形或蛆狀,剛孵化的一齡幼蟲體表具有形態和數目固定的剛毛,稱為一次刺毛,對分類與演化研究很有價值。其後每蛻一次皮即稱為增加一齡,而體表會長出變化多端,數目不定的二次刺毛。幼蟲期總齡數多為四或五齡,也有多達12齡的情形。 幼蟲頭部骨化程度高,生有感覺毛,下端具備咀嚼式口器,兩側各有數個可用來感光的側單眼。頭部具有縫線,藉以在化蛹時裂開讓蛹體脫出。 幼蟲身軀體壁骨化程度弱而呈膜質,骨化部分只有一次刺毛基部的小硬板前胸背側稱為前胸硬皮板的部分,以及第10腹節背側稱為肛上板的部分。 胸部具有三對步足,其跗節僅一節,末端有一爪。腹部分為10節,於第3、4、5、6及第10節具有原足。原足上具有成列小鉤,稱為原足鉤,藉以附著攀附於幼蟲吐絲形成的絲墊或其他物體上[14]。前胸以及第1至8腹節各具一對氣孔。幼蟲多半總齡數固定,但不固定的種類也不少。
蝴蝶的蛹屬於彼蛹,附肢與翅緊貼軀體,體表在不同部位生有刺毛。頭部的複眼,觸角及小顎外葉等部分可見。觸角沿前翅前緣延伸小顎外葉位於腹面中央,其長度常超出翅面。 胸部各節均可見,多以中胸最為發達。前翅位於蛹體側腹面,後翅大部分或完全為前翅覆蓋。前足,中足可見,位於觸角與小顎外葉之間。中胸氣孔位於背側。腹部10節均可見,但最後三節癒合而不可動其他腹部的可動性依種類而不同。第1至8腹節具有氣孔,但是第1腹節氣孔因被翅覆蓋而不可見,第8腹節氣孔則失去功能而無開口。交尾器痕在雄蟲見於第9腹節,而在雌蟲則在第8及9腹節,交尾器痕呈圓形或線形。肛門痕位於第10腹節,形狀也呈圓形或線形。 第10腹節特化,伸長成稱為垂懸器的構造,有鉤狀刺毛,蛹體可藉此構造攀附絲上以附著於物體上蛹體附著物體的方式主要分為僅以垂懸器附著,頭下尾上的懸蛹以及胸部有絲帶環繞固定的縊蛹,不過也有許多屬於以上兩類的其他附著方式。蛹的外部形態變化萬端,許多種類模擬植物體不同部位藉以隱藏有些種類並其依附著位置的質地,色彩而呈綠色或褐色。另有一些體內含有毒素的種類擁有鮮豔色彩。
習性

- 食性
蝴蝶成蟲的口器特化為吸管狀,所以只能進食液態的食物。除了花蜜以外,水、樹汁、腐果、動物糞便等都是各種蝴蝶的食物。蝴蝶喜歡在溪邊或濕潤的土壤聚集一起吸水,攝取水中的礦物質。

- 共生
許多灰蝶幼蟲會與螞蟻共生,螞蟻保護幼蟲,並從幼蟲身上取得蜜露,雙方都獲得好處[15]。但蟻種不限定,沒螞蟻時幼蟲亦可生長,此爲非絕對的互利共生;銀線灰蝶屬幼蟲亦與螞蟻共生,但雌性蝴蝶要有特定蟻種氣味時才會產卵,這是絶對的互利共生。
- 過冬

冬天的時候蜜源植物不多,寄主植物也凋零,成蟲和幼蟲會因食物不足而難以生存。所以很多蝴蝶都以卵或蛹形態過冬,因為這兩個都是靜止狀態,不需進食。然而,有些斑蝶會以成蟲型態過冬,在秋季牠們會努力吸花蜜,儲存養分,冬季時會集體在山谷中過冬,形成蝴蝶谷的奇觀。帝王斑蝶和大絹斑蝶兩種較爲出名的種都會進行長距離遷徙。
- 領域
雄性蝴蝶的領域行爲會驅離其他競爭者,讓自身取得制空優勢。牠們會站在樹上的葉面以取得制空權,假如有其他雄蝶或飛行物闖入領域,便會互相追逐,直至入侵者被驅逐。但如果闖入的是雌蝶,便會設法將雌蝶留住並交配。領域性是蝴蝶常見的競爭方式。
- 登峰
在每天的上午,各地的蝴蝶會順着地形往上飛,最後聚集在各處山嶺或山脊。雄蝶在山頭盤據,互相追趕,等待雌蝶登山。而雌蝶在交配後會飛到山下產卵。
- 求偶
雄蝶的數量比雌蝶多,所以雄蝶的競爭非常激烈,數隻雄蝶追求一隻雌蝶的情況並不罕見。雄蝶對雌蝶的氣味異常敏感,常有雄蝶守候在未羽化的雌蝶蛹旁,待雌蝶羽化後便第一時間交配。雄蝶的領域和登峰行為都是為了交配繁殖。
生態

天敵
蝴蝶位於食物網的底層,有多种天敵,是雀鳥、蜥蜴、蜘蛛、螳螂、蜻蜓、青蛙等多种生物的捕食对象。牠們大多行動敏捷或會設置陷阱,会捕捉正在訪花甚至飛行中的蝴蝶成蟲。蝴蝶幼生期的各階段亦幾乎都有寄生性天敵,如小自卵蜂、繭蜂、大至姬蜂、寄生蠅、線蟲等。不同种类的天敌會选择把卵產在蝴蝶的卵、幼蟲或蛹上。當牠們的幼蟲孵出後就會以寄主的組織為食。這類寄生生物吸取寄主營養長大後通常會導致寄主死亡,這類關係屬擬寄生物,擬寄生生物與寄主之間大多具專性。此外,細菌、病毒和真菌都是令牠們生病的微生物。
防禦

部分灰蝶幼蟲與螞蟻共生時,能藉由螞蟻提供的防衛能力避免被擬寄生生物危害,但遇上體型稍大的天敵,螞蟻們也使不上力。幼蟲及成蝶可藉由移動能力尋找合適地點躲藏,或是快速飛行,逃跑或裝死,而沒有移動能力的蝶卵,蛹,則有其他避敵方法提高生存機會。
- 鱗片
蝴蝶身上佈滿細毛和鱗片,當天敵捕食時,容易脫落的細毛和鱗片令敌人不能抓緊,增加逃脫的機會。
- 隱藏
蝴蝶會藉由各種隱蔽環境隱藏起來。例如臺灣灑灰蝶雌蝶將卵產在樹皮裂縫內;弄蝶幼蟲製作的蟲巢,進食時間外到都會在葉包內;趾弄蝶屬成蝶停棲在葉片下表面等。
.jpg.webp)
- 僞裝
蝴蝶的各種外型、顏色或行爲等融入周遭環境,利用保護色讓天敵不易察覺,例如很多蝴蝶的幼蟲和卵到和寄主植物的顏色十分相似,偽裝成葉片、花或樹枝的一部份,好像枯葉蛺蝶和蠹葉蛺蝶的翅紋像枯葉[16]。又會模仿天敵不感興趣的物體,如鳳蝶幼蟲外型像鳥糞[17]。又會利用斑紋的形態分割干擾天敵的視覺,如環蛺蝶屬翅膀的條紋破碎、灰蝶後翅臀角的眼斑、尾突,讓天敵以爲是頭部而攻擊錯誤目標,所以一些灰蝶停留時會擺動後翅來模仿頭部動作,天敵往往被騙以攻擊後翅,灰蝶藉此逃過一劫[18]。

- 威嚇防衛
眼蝶等蝶種翅膀擁有不合比例的巨大眼斑或鳳蝶屬終齡幼蟲胸部的眼斑等,都可能讓天敵誤以爲是巨獸而不敢攻擊。大量的幼蟲群聚也有威嚇效果,亦令捕食者難以集中捕食,令其生存機會增加[19]。部分蝶蛹表面具有金屬色澤,斑蝶幼蟲顏色鮮豔,對比明顯的體色屬於警戒色,警告天敵不要捕食。部分蝴蝶幼蟲體表有堅硬棘刺或是體表有長毛,藉以警告天敵牠們可能有毒,不可口或難以下嚥。若無法嚇阻天敵的行動,鳳蝶幼蟲的臭角散發難聞的氣味[20]和斑蝶成蝶會用毛筆器散發出斑蝶素的味道以達到自我防衛的驅敵效果。
- 擬態
擬態是屬於利用其他生物的防衛機制,擬態要有以下三者才成立:捕食者、被擬態者及擬態者。捕食者在捕食過程中學會被擬態者不好吃有毒或難以捕食,由不好的經驗讓牠之後會避免捕食外型相似的物種,擬態者則因爲外型像被擬態者而減少被捕食。
- 貝氏擬態:無威脅性、無毒者,模仿具威脅性、有毒的外貌,猶如狐假虎威[19]。被擬態者的數量要夠多且兩者要共域分布才符合,例如雌性金斑蛺蝶外型像有毒的虎斑蝶,玉帶鳳蝶雌蝶紅紋型像有毒的紅珠鳳蝶。
- 穆氏擬態:兩種或兩種以上都是具威脅性、有毒的物種,彼此外型相似互爲擬態者與被擬態者。體色通常爲鮮豔或對比色,擬態能加強警戒色的效果例如以馬兜鈴爲食的鳳蝶多具有白色斑紋或紅色斑紋;青斑蝶屬、絹斑蝶屬、旖斑蝶屬三類有毒的成蝶的外型相似,有毒的紫斑蝶屬中的物種有相似的色澤等。
- 速度擬態:速度慢者模仿速度快者的樣貌以欺騙天敵,做出放糜追捕的念頭。
- 隱蔽式擬態:獵物(擬態者)與生活的環境或周遭物體相像(被擬態者),藉此躲避天敵的捕食。此作法乃僞裝模仿等納入的廣義擬態。
演化
鱗翅目的兩個亞目
最傳統的鱗翅目分類只是依物種的觸角形態把蝴蝶和蛾區分,分類名稱分別為「錘角亞目」和「異角亞目」。
| 鱗翅目 |
| ||||||
Kawahara (2014)
蝴蝶是鱗翅目中的一個總科級演化支,稱為鳳蝶總科(Papilionoidea)。過往鱗翅目曾依據觸角形態分為錘角亞目(Rhopalocera,蝶類)和異角亞目(Heterocera,蛾類),但現今研究已證實異角亞目為併系群,這種分類並不合宜。其後,一些分子種系發生學研究分別有指出蝴蝶跟小鱗翅亞目或大鱗翅亞目有密切的親緣關係[22][23]。近年的種系發生學研究都肯定鳳蝶總科是鱗翅目之中,雙孔亞目(Ditrysia)之中、短突雙孔類(Apoditrysia)之中、螟蛾類(Obtectomera)之中的一個演化支,然而對於鳳蝶總科與鄰近蛾類之親緣關係的層面上,在各個研究結果中都不時發生變動。較新的研究分析如下[2]:
| 螟蛾類 Obtectomera |
| ||||||||||||||||||||||||||||||||||||||||||||||||
M. J. Scoble (1992)
- M. J. Scoble (1992) 的分類法把蝴蝶分成3個總科,6科。是首先把喜蝶歸入蝶類。另外,當時蜆蝶歸入灰蝶科中。[25]
| ||||||||||||||||||||||||||||
周堯 (2006)
- 周堯 (2006) 的分類法把蝴蝶分成4個總科,17科。[26]
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Heikkilä (2015)
在過往的鱗翅目分類系統中,蝴蝶所屬的錘角亞目曾經分為3個總科:弄蝶總科、鳳蝶總科和喜蝶總科[25],並認為弄蝶是較原始的類群。然而,近年的分子種系發生學研究結果都否定這種分類法。較新的研究數據發現鳳蝶的分類才是蝴蝶演化支的基群,即鳳蝶科是所有其他蝶類(包括弄蝶和喜蝶)所構成之演化支的姐妹群,換言之,以前鳳蝶總科和弄蝶總科的定義也成為了多系群[27],因此蝴蝶不應區分成三個總科[28]。綜觀而言,現今蝴蝶共分類為1個總科,當中包含了7個科。各科之間的關係為:蜆蝶科和灰蝶科是姐妹群,她們的共同演化支是蛺蝶科的姐妹群,粉蝶科是以上的姐妹群,弄蝶科+喜蝶科的共同演化支是以上的姐妹群,最後鳳蝶科是以上各科的姐妹群[1]。
| 鳳蝶總科 |
| ||||||||||||||||||||||||||||||||||||
圖集






.jpg.webp)
文化

- 大型蝴蝶非常引人注意,有人专门收集各种蝴蝶标本,亦吸引许多自然愛好者和攝影者參與
- 愛爾蘭的傳統民族相信人死後會化成蝴蝶[29]
- 中国哲學家莊周曾經夢見自己變為蝴蝶,是為莊周夢蝶,认为人不可能确切区分真实和虚幻
- 中国传说梁山伯与祝英台当中,男女主人双双殉情后,灵魂化为蝴蝶。蝴蝶在当中成为追求自由与爱情的象征
- 希臘羅馬神話故事裏,也有被描繪成背上生著一對蝴蝶翅膀的美女─普賽克(Psyche)
- 蝴蝶的閩語泉州話為尾蝶[30]
- 觀蝶
觀蝶就是觀察蝴蝶自然生態的活動,可以到郊外或蝴蝶園進行。觀蝶時不應搔擾野生動物、採捕蝴蝶的任何階段,以免擾亂生態。觀蝶時可攜帶當地的蝴蝶圖鑑、記錄表和相機等工具,方便作記錄[9]。
參見
腳註
- Chazot, N., Wahlberg, N., Freitas, A. V. L., Mitter, C., Labandeira, C., Sohn, J. C., ... & Heikkilä, M. (2019). Priors and Posteriors in Bayesian Timing of Divergence Analyses: the Age of Butterflies Revisited. Systematic biology. https://doi.org/10.1093/sysbio/syz002
- Heikkilä, M., Mutanen, M., Wahlberg, N., Sihvonen, P., & Kaila, L. (2015). Elusive ditrysian phylogeny: an account of combining systematized morphology with molecular data (Lepidoptera). BMC evolutionary biology, 15(1), 260.
- Mitter, C., Davis, D. R., & Cummings, M. P. (2017). Phylogeny and Evolution of Lepidoptera. Annual review of entomology, 62, 265-283.
- Dönitz, Prof. Dr. W. . Berliner entomologische Zeitschrift. 1897, 42 (1-2): 1–25. doi:10.1002/mmnd.18970420102.(德文)
- Condamine, F. L., Toussaint, E. F., Clamens, A. L., Genson, G., Sperling, F. A., & Kergoat, G. J. (2015). Deciphering the evolution of birdwing butterflies 150 years after Alfred Russel Wallace. Scientific Reports, 5, 11860.
- 大衛.卡特. . 中國友誼出版公司. 2007年7月. ISBN 9787505713222.
- Williams, Ernest; Adams, James; Snyder, John. . The Lepidopterists' Society. [9 September 2015]. (原始内容存档于2015年5月13日).
- Venkatesha, M. G.; Shashikumar, L.; Gayathri Devi, S.S. . Current Science. 2004, 87 (5): 571–572.
- 潘瑞輝 譚健聰. . 香港: 大埔環保會 鳳園蝴蝶保育區. 2011年8月. ISBN 978-988-15271-1-0.
- Smart, Paul 1977. The Illustrated Encyclopedia of the Butterfly World, chapter 2. Chartwell Books. ISBN 0-89009-093-9
- Triplehorn, Charles A. and Johnson, Norman F. 2005. Borror and Delong's introduction to the study of insects. 7th ed, Thomson Brooks/Cole. Belmont, California, p578. ISBN 0-03-096835-6
- Ball, P. (2012). Nature's color tricks. Scientific American, 306(5), 74-79.
- Monteiro, A.; Pierce, N. E. . Molecular Phylogenetics and Evolution. 2001, 18 (2): 264–281. PMID 11161761. doi:10.1006/mpev.2000.0872.
- . Larva Legs. Chicago Academy of Sciences. [7 June 2012]. (原始内容存档于2012-03-19).
- Fiedler, K.; Holldobler, B.; Seufert, P. . Cellular and Molecular Life Sciences. 1996, 52: 14–24. doi:10.1007/bf01922410.
- Robbins, Robert K. . American Naturalist. 1981, 118 (5): 770–775. doi:10.1086/283868.
- . University of Florida. [12 September 2015]. (原始内容存档于2019-06-11).
- Stevens, M. . Biological Reviews. 2005, 80 (4): 573–588. PMID 16221330. doi:10.1017/S1464793105006810.
- Edmunds, M. . Longman. 1974: 74–78, 100–113.
- Chattopadhyay, J. 2011. The structure and defensive efficacy of glandular secretion of the larval osmeterium in Graphium agamemnon agamemnon Linnaeus, 1758 (Lepidoptera: Papilionidae).
- Kawahara, A. Y., & Breinholt, J. W. (2014, August). Phylogenomics provides strong evidence for relationships of butterflies and moths. In Proc. R. Soc. B (Vol. 281, No. 1788, p. 20140970). The Royal Society.
- Minet J. 1991 Tentative reconstruction of the ditrysian phylogeny (Lepidoptera, Glossata). Entomol. Scand. 22, 69–95. (doi:10.1163/187631291X00327 (页面存档备份,存于))
- Tree of Life Web Project. 2010. Ditrysia (页面存档备份,存于). Version 17 November 2010 (temporary). in The Tree of Life Web Project (页面存档备份,存于) (英文)
- van Nieukerken et al., 2011 in Zhang (Ed.), Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Order Lepidoptera Linnaeus, 1758 Zootaxa 3148 : 212-221
- Scoble MJ. 1992 The Lepidoptera. Form, function and diversity. Oxford, UK: Oxford University Press.
- 寿建新, 周尧, 李宇飞. . 中國: 陝西科學技術出版社. 2006-04-01. ISBN 9787536936768.
- Kristensen NPAckery PR, de Jong R, Vane-Wright RI. 1998 The butterflies: Hedyloidea, Hesperoidea and Papilionoidea. In Lepidoptera: moths and butterflies 1 Handbuch der Zoologie/handbook of zoology IV/35 (ed. Kristensen NP), pp. 263–300. Berlin, Germany: Walter de Gruyter.
- Kristensen NPKristensen NP, Skalski AW. 1998 Phylogeny and palaeontology. In Lepidoptera: moths and butterflies 1 Handbuch der Zoologie/handbook of zoology IV/35 (ed. Kristensen NP), pp. 7–25. Berlin, Germany: Walter de Gruyter.
- 寄藤文平, 梁桂慈譯. 2010. 死的型錄:鬼才插畫家筆下的生命終點. 圓神出版社有限公司
- 蒲实. . 《三联生活周刊》. 2020年12月21日, 1118 (51) [2021-07-01]. (原始内容存档于2021-07-01).
參考
| 维基共享资源中相關的多媒體資源:蝴蝶(分類) |
| 維基物種上的相關:蝴蝶 |
| 维基词典中的词条「」。 |
- 中國
- 武春生、徐堉峯. . 海峡书局. 2017. ISBN 9787556703029 (中文(简体)).
- 周堯. . 中國: 河南技術出版社. 1992年. ISBN 75349-15740.
- 寿建新, 周尧, 李宇飞. . 中國: 陝西科學技術出版社. 2006-04-01. ISBN 9787536936768.
- 台灣
- 呂至堅, 陳建仁. . 台灣: 晨星出版有限公司. 2014年3月23日. ISBN 9789861777849.
- 徐堉峰. . 台灣: 晨星出版有限公司. 2013年2月10日. ISBN 9789861776705.
- 徐堉峰. . 台灣: 晨星出版有限公司. 2013年2月10日. ISBN 9789861776705.
- 徐堉峰. . 台灣: 晨星出版有限公司. 2013年3月10日. ISBN 9789861776682.
- 香港
- 潘瑞輝. . 環保協進會. 2020. ISBN 9789881527295.
- 李東霖、吳滄桑、丘紹文、楊建業. . 香港鱗翅目學會. 2020. ISBN 9789881220905.
- 楊建業、吳滄桑、王怡君. . 香港鱗翅目學會. 2015. ISBN 9789881220967.
- 潘瑞輝、楊英豪. . 香港鱗翅目學會. 2012. ISBN 978-962-86943-7-2.
- 其他
- Löfstedt, C., Wahlberg, N. & Millar, J. G. 2016: Evolutionary Patterns of Pheromone Diversity in Lepidoptera. - In: Allison, J.D. & Cardé, R.T. (eds.), pp. 43–78, Pheromone Communication in Moths: Evolution, Behavior and Application. University of California Press, Berkeley. (英文)
外部連結
- 台灣蝴蝶保育學會
- 香港市區公園的蝴蝶 - 品種和分類 (页面存档备份,存于) - 康樂及文化事務署
- Papilionoidea (页面存档备份,存于) - funet.fi