有孔蟲門
有孔蟲門(学名:),為變形蟲狀原生生物的大分類。牠們擁有的網狀假足及幼細線狀細胞質會分散及融合而形成動態的網[2],它們會形成有一個或多個室的外殼,部分在結構上有高度發展[3]。牠們大部分大小小於一毫米,但部分則較大,紀錄中最大的樣本大小達19厘米[4]。
| 有孔虫门 化石时期: | |
|---|---|
 | |
| 《自然界的藝術形態》 (1904), 圖版 2: Thalamphora | |
 | |
| 《自然界的藝術形態》 (1904), 圖版 81: Thalamophora | |
| 科学分类 | |
| 演化支: | SAR 超类群 SAR |
| (未分级): | 有孔蟲類 Rhizaria |
| 总门: | 有孔虫总门 Retaria |
| 门: | 有孔虫门 Foraminifera d'Orbigny, 1826 |
| 綱與目 | |
雖然現尚未有形態學上相關的支持,分子資料強力提出因為有孔蟲門與絲足蟲類()及放射蟲門()都為變形蟲狀及有複雜形態的外殼,所以有密切關係。這三類形成有孔蟲界()[5]。但有孔蟲門與其他類別的實際關係還不是完全清楚。
分類
截至2020年1月8日,WoRMS紀錄有孔蟲門物種可以分為以下五大綱[6]:
- †紡錘蟲綱 Fusulinata Fursenko, 1958 [7][8]
- 非紡錘形亞綱 Afusulinana Vachard, Pille & Gaillot, 2010 †[9][10][11]
- Order Archaediscida †
- Order Earlandiida †
- Order Parathuramminida †
- Order Pseudopalmulida †
- 紡錘形亞綱 Fusulinana †[12][10]
- Order Endothyrida †
- 紡錘蟲目 Fusulinida †
- 非紡錘形亞綱 Afusulinana Vachard, Pille & Gaillot, 2010 †[9][10][11]
- Globothalamea
- Carterinida
- Lituolida
- Acupeinidae
- Adercotrymidae
- Ammobaculinidae
- Ammolagenidae
- Ammosphaeroidinidae
- Aschemocellidae
- Barkerinidae
- Conotrochamminidae
- Debarinidae
- Discamminidae
- Duquepsammiidae
- Duquepsamminiidae
- Dusenburyinidae
- Glaucoamminidae
- Haplophragmoididae
- Hormosinellidae
- Kunklerinidae
- Lituolidae
- Lituotubidae
- Marieitidae
- Mayncinidae
- Nautiloculinidae
- Nezzazatidae
- Nouriidae
- Oxinoxisidae
- Pavonitinidae
- Piallinidae
- Placopsilinidae
- Plectorecurvoididae
- Plectoverneuilinellidae
- Plectoverneuilinellidea
- Ponceamminidae
- Prolixoplectidae
- Pseudobolivinidae
- Remaneicidae
- Reophacellidae
- Reophacidae
- Rzehakinidae
- Sphaeramminidae
- Spiroplectamminidae
- Textulariopsidae
- Trilocularenidae
- Tritaxiidae
- Trochamminidae
- Trochamminoidae
- Verneuilinidae
- Loftusiida
- Robertinida
- Rotaliida
- Textulariida
- Monothalamea
- Nodosariata
- Tubothalamea
- 其他地位未明分類單元
這五個綱還包括以下各目:
- 網足蟲亞目 Allogromiida
- Carterinida
- †紡錘蟲目 Fusulinida
- 抱球蟲目 Globigerinida
- †Involutinida
- Lagenida
- 粟孔蟲 Miliolid
- Robertinida
- 輪蟲目 Rotaliida
- Silicoloculinida
- Spirillinida
- Textulariida
- 地位未定
- Xenophyophorea
- Reticulomyxa
现生有孔蟲門生物
生存環境及形態

現代有孔蟲門生物主要為海洋生物,它們也可以在其他含有一定盐分的環境生存[13]。部分物種在淡水生存及一種在潮濕的雨林土壤中生活。它們在小型底棲生物(meiobenthos)中十分常見,有約40種為浮游生物[2]。但是,这个数字可能只部分正确,因為有很多基因上不同的物種在形態上是不能夠区分的[14]。其細胞分為粒狀內質(endoplasm)及透明外質(ectoplasm)。其假足狀的網可以在一個開口或在外殼的很多穿孔中出現,其特色是當中有細粒在網中自由流動[13]。
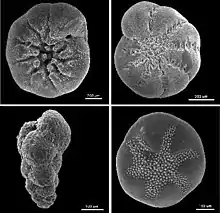
外殼
成分
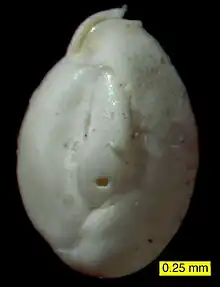
外殼的形狀及成分是对有孔蟲門生物进行分类和鉴定的主要方法。大部分有孔虫们生物具有钙质(石灰质)外殼,由碳酸鈣形成[13]。或为有機質,或由有机物或钙质成分粘接細小的沉積物形成,硅质外壳极为罕见。
演化學上的重要性
化石的發現
死去的浮游有孔蟲門生物不斷大量地向海床落下,它們蘊含豐富礦物的外殼以化石的形態在沉積物中被保留著。在1960年代起,一方面在深海鑽探計劃(Deep Sea Drilling Program)、海洋鑽探計劃(Ocean Drilling Program)及綜合海洋鑽探計劃(Integrated Ocean Drilling Program)的贊助,另一方面亦受到石油探採(oil exploration)的目的所推動下,由先進的深海鑽探技術所帶出的沉積物核心中,可以發現幾百萬年前的有孔蟲門生物化石。
大量的高品質素材
在其實際上無限量供應的化石外殼及其核心中有相對高準確度的時期抑制原型下,有孔蟲門生物能夠製造出非常高品質的浮游有孔蟲門生物化石記錄,最早可以追溯至侏羅紀中期,其化石亦可以提供科學家一個不平衡的記錄去測試及記錄演化過程。因為化石記錄的品質極高,令物種間的相互關係可以以此為基礎,能夠造成極詳細的影像。在很多情況下其後會在現存的樣本上以分子基因研究的方法作獨立確認。
有孔蟲門生物的應用
地層測量
因為有孔蟲門生物的多樣性、數量充足及複雜的形態的關係,有孔蟲門生物的化石集合物對生物地層學(biostratigraphy)十分有用,亦可以用作準確地測量出岩石的年代。石油工業(oil industry)十分依賴微化石(microfossil)如有孔蟲門生物去找尋潛在石油庫存。
參考資料
- Parfrey, Laura Wegener; Lahr, Daniel J. G.; Knoll, Andrew H.; Katz, Laura A. . Proceedings of the National Academy of Sciences of the United States of America. August 16, 2011, 108 (33): 13624–13629 [2020-01-13]. Bibcode:2011PNAS..10813624P. PMC 3158185
 . PMID 21810989. doi:10.1073/pnas.1110633108. (原始内容存档于2019-02-18).
. PMID 21810989. doi:10.1073/pnas.1110633108. (原始内容存档于2019-02-18). - Hemleben, C.; Spindler, M.& Anderson, O.R. . Springer-Verlag. 1989: 363.
- Kennett, J.P.; Srinivasan, M.S. . Hutchinson Ross. 1983: 265.
- Pavlovec, R. . Proteus. 1987, 49 (5): 167–169.
- Cavalier-Smith, T. . European Journal of Protistology. 2003, 34 (4): 338–348.
- Gordon, Dennis; Hayward, Bruce W. . WoRMS. 2011-10-17 [2020-01-08].
- Hayward, Bruce W. . WoRMS. 2017-12-08 [2020-01-08].
- Fursenko, A. V. [Fundamental state of development of foraminiferal faunas in the geological past]. ? (Minsk: Trudy Instituta Geologicheskikh Nauk, Akademiia Nauk Belorusskoi SSR). 1958, 1: 10–29.
- Le Coze, François; Hayward, Bruce W. . WoRMS. 2019-07-06 [2020-01-08].
- Vachard, D.; Pille, L.; Gaillot, J. . Revue de Micropaléontologie. 2010, 53 (4): 209–254 (英语).
- Gaillot, J.; Vachard, D. . Coloquios de Paleontologia. 2007, 57: 37–223 [2020-01-08]. (原始内容存档于2020-04-22) (英语).
- LeCoze, François; Hayward, Bruce W. . WoRMS. 2019-07-06 [2020-01-08].
- Sen Gupta, B.K. . Springer. 1983: 384.
- Kucera, M.; Darling, K.F. . Philosophical Transactions of the Royal Society of London. 2002, A360 (4): 695–718.
- Bernhard, J. M.; Bowser, S.M. . Earth Science Reviews. 1999, 46: 149–165.
- Foraminifera: History of Study (页面存档备份,存于), University College London, retrieved 20 September 2007
- Langer, M. R.; Silk, M. T. B., Lipps, J. H. . Journal of Foraminiferal Research. 1997, 27 (4): 271–277. 外部链接存在于
|title=(帮助) - Adl, S. M.; Simpson, A. G. B., Farmer, M. A., Anderson, R. A., Anderson, O. R., Barta, J. A., Bowser, S. M., Brugerolle, G., Fensome, R. A., Fredericq, S., James, T. Y., Karpov, S., Kugrens, P., Krug, J., Lane, C. E., Lewis, L. A., Lodge, J., Lynn, D. H., Mann, D. G., McCourt, R. M., Mendoza, L., Moestrup, O., Mozley-Standridge, S. E., Nerad, T. A., Shearer, C. A., Smirnov, A. E., Speigel, F. W., Taylor, M. F. J. R. . Journal of Eukaryotic Microbiology. 2005, 52 (5): 399–451.
- Zachos, J.C.; Pagani, M., Sloan, L., Thomas, E., and Billups, K. . Science. 2001, 292: 686–693.
- . [2008-03-01]. (原始内容存档于2019-09-16).
外部連結
- The University of California Museum of Paleontology website has an Introduction to the Foraminifera (页面存档备份,存于)
- The star*sand project(part of micro*scope)is a cooperative database of information about foraminifera
- The Smithsonian Institution's National Museum of Natural History has the largest type collection of foraminifera in the world
- Researchers at the University of South Florida developed a system using foraminifera for monitoring coral reef environments (页面存档备份,存于)
- University College London's micropaleontology site (页面存档备份,存于) has an overview of foraminifera, including many high-quality SEMs
- Illustrated glossary of terms used in foraminiferal research (页面存档备份,存于) is the Lukas Hottinger's glossary published in the OA e-journal "Carnets de Géologie - Notebooks on Geology" (页面存档备份,存于)
- CHRONOS has several foraminifera resources,including a taxon search page and a micro-paleo section
- eForams is a web site focused on foraminifera and modeling of foraminiferal shells
- Benthic foraminifera information (页面存档备份,存于) from the 2005 Urbino Summer School of Paleoclimatology
- Information on Foraminifera Martin Langer's Micropaleontology Page
- 有孔虫画廊-插图目录 与5000+影像