蛇
蛇是一类無足的爬蟲類動物,是蛇亚目(学名:)的通称,属于爬行纲,是蜥蜴的一個演化支,另有其它無足的爬虫類如蚓蜥、蛇蜥等并不属于蛇亚目。
| 蛇|蛇亚目 | |
|---|---|
 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | Reptilia |
| 目: | 有鳞目 Squamata |
| 演化支: | 有毒类 Toxicofera |
| 演化支: | 蠎形类 Pythonomorpha |
| 演化支: | 蛇形类 Ophidiomorpha |
| 演化支: | 泛蛇类 Ophidia |
| 亚目: | 蛇亚目 Serpentes Linnaeus,1758 |
| 下目 | |
| |
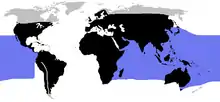 | |
| 藍色:海生蛇类;黑色:陆生蛇类 | |
又有虵、虺、螣、蚦、蜧、蜦、长虫等別稱,根據種類也會有蟒、蚺、蝰、蝮等近義稱呼。正如所有爬蟲類有鱗目一樣,蛇類全身佈滿鱗片。所有蛇類都是肉食性動物。目前全球共有3,000多種蛇類,包括體型最短小的細盲蛇科以至最長的蟒科及蚺科。為了配合蛇類窄長的身體,成對的內臟(如肺、腎)會在蛇體前後排列,而非左右互對。
部分蛇類擁有毒性,能使被其咬擊的生物受傷、疼痛以至死亡。蛇的另一個特徵是顎部能作出廣角度的開合,因此能吞食比自己身型龐大的獵物。生物研究指蛇類大概於白堊紀時代由蜥蜴類衍生而成。現代蛇類的分類研究,大概可追溯至古新世時代。目前紀錄中最巨型的蛇類是活於古新世的泰坦巨蟒,長度達13米長,其化石被發現的年份是2009年[1];目前體型最細小的蛇類是卡拉細盲蛇,長度約只有10公分[2]。
分類
蛇類主要分成兩大類,分別是真蛇下目及盲蛇下目。
真蛇下目:共15個科 | ||||
| 種類 | 學名/命名者/命名年 | 例種 | 圖片 | |
|---|---|---|---|---|
| 瘰鱗蛇科 | Acrochordidae Bonaparte,1831[3] | 瘰鱗蛇 | ||
| 筒蛇科 | Aniliidae Stejneger,1907[4] | 紅尾筒蛇 | ||
| 倭管蛇科 | Anomochilidae Cundall、Wallach及Rossman,1993[5] | 李奧拿氏倭管蛇 | ||
| 穴蝰科 | Atractaspididae Günther,1858[6] | 穴蝰 | ||
| 蚺科 | Boidae Gray,1825[3] | 森蚺 |  | |
| 島蚺科 | Bolyeriidae Hoffstetter,1946 | 島蚺 | ||
| 游蛇科 | Colubridae Oppel,1811[3] | 食卵蛇、瘦蛇 |  | |
| 管蛇科 | Cylindrophiidae Fitzinger,1843 | 紅尾管蛇 | ||
| 眼鏡蛇科 | Elapidae Boie,1827[3] | 眼鏡蛇、眼鏡王蛇、印度眼鏡蛇、虎蛇、死亡蛇 |  | |
| 美洲閃鱗蛇科 | Loxocemidae Cope,1861 | 美洲閃鱗蛇 |  | |
| 蟒蛇科 | Pythonidae Fitzinger,1826 | 緬甸蟒、盾蟒、網紋蟒、窩瑪蟒 |  | |
| 林蚺科 | Tropidophiidae Brongersma,1951 | 古巴林蚺 | ||
| 盾尾蛇科 | Uropeltidae Müller,1832 | 紅腹銼尾蛇 |  | |
| 蝰蛇科 | Viperidae Oppel,1811[3] | 角蝰 |  | |
| 閃鱗蛇科 | Xenopeltidae Bonaparte,1845 | 海南閃鱗蛇 |  | |
盲蛇下目:共3個科 | ||||
| 種類 | 學名/命名者/命名年 | 例種 | 圖片 | |
| 異盾盲蛇科 | Anomalepidae Taylor,1939[3] | 特立尼達盲蛇 | ||
| 細盲蛇科 | Leptotyphlopidae Stejneger,1892[3] | 德州細盲蛇 |  | |
| 盲蛇科 | Typhlopidae Merrem,1820[7] | 鉤盲蛇 |  | |
演化
關於蛇類的種系發生學資料是相當貧乏的,這是因為蛇的骨骼較為細小及易碎,難以保存,能夠成形的完整化石更是並不常見,研究材料顯然不足。不過,一些在南美洲及非洲發現的蛇類化石,其骨骼構造與蜥蜴相近,讓科學家可以確定及辨認那些經歷了一億年的蛇類。[8]從形態學的基礎而言,學界已普遍同意蛇類是系出於蜥蜴類的。[8][9]從兩者分子結構研究所得的結果,更充分地支持這個說法;目前學者假設擁有毒素的蛇類是遠古毒蜥蜴類生物的毒素直接繼承者(詳見蛇毒)。
從蛇類化石的研究中,學者認為蛇類是白堊紀時代的洞棲蜥蜴一系演化而成的,例如巨蜥或其它鄰近種類[10]。一條早期被發現並被命名為「Najash rionegrina」的蛇類化石,相信是以雙足活動的洞棲性生物,牠的身體已經擁有薦骨,而且其生活習性完全是陸行性的[11]。一種被認為與上述化石相似的現存生物,是位於東南亞婆羅洲的拟毒蜥,不過這種巨蜥是半水行性的,與前面所說的古蛇化石的習性仍有不同之處[12]。由於這些遠古生物是生活於地洞中的,牠們的四肢逐漸退化消失,其身體亦變得更強調流線型,方便進出於地穴之間[12]。根據這個假設,蛇類的一些特徵如其擁有透明的眼膜(功能類似眼瞼)、欠缺外耳等,都能解釋是為了方便牠們在地穴行動的特性而演化成的[10][12]。另外還有一些上古時代的蛇類是擁有後肢的,但牠們身體的盆骨與脊椎之間卻沒有直接的聯繫,這類蛇包括厚針龍及真足蛇等,其存活歷史相信比「Najash」化石更為久遠[13]。


在現代蛇類中較為原始的蛇種中,蟒蛇及蚺蛇還保存著已經退化的後肢痕跡,牠們身體上殘餘一雙巨大呈爪狀的幼肢,這雙爪子現在的功能是充當雄蛇在交配時藉以抓緊雌蛇身體的工具[8][13]。盲蛇科及細盲蛇科是蛇類中至今仍保留有盆骨帶的零星例子,這段盆骨帶有時更會從體內突出,可以直接觀察得到。
基於同源基因的變化,所有蛇類的前肢都已經不再存在。蛇類的中軸骨骼構造(脊椎結構)與一般的四肢動物無異,根據身體的段落而能分為不同的區域,包括頸椎、胸椎、腰椎、盆椎及尾椎等。中軸骨骼生物的同源異型基因強化了蛇類的胸椎,而牠們曾經存在過的前肢亦被胸椎發展所同化,極端發展的胸椎更進而組成了大部分蛇類的主要骨骼結構。蛇的胸椎骨外長有肋骨,而其頸椎、腰椎及盆椎的節數則大幅減少(目前只剩下2至10節腰椎及盆椎),其短小的尾部亦只由餘下的少量尾椎所構成,不過仍有多蛇種能善用牠們的尾部,尤其對於一些水行性及樹棲性的蛇類而言。另外毒蛇的牙都是空心的,牠們其中一種攻擊模式是先以身體捲住獵物,然後用毒牙咬著對方,將毒汁注入獵物體內。蛇毒多以蛋白質為主,根據各種蛇毒性質的分別來破壞獵物的神經系統或其它生物系統,從而令對方死亡。

另一種基於生物形態學的假設認為,蛇類及巨蜥類的祖先其實是滄龍:一種存在於白堊紀並已經滅絕的水行性爬蟲類生物[9]。在這種假設下,蛇類的透明眼膜將會被解釋成是為了便於在水中行動因而演變成的,而失去外耳的原因是因為外耳在水中本來就不能發揮功能,這些演變的最終型態便是今天的海蛇。在白堊紀後期,蛇類重新返回陸地上生活,從而發展成今天的各種陸行型蛇類。在一些殘餘的蛇類化石中能找到白堊紀時期的海水沉澱物,更能確認以上的假設,因此亦可判斷出這批化石比陸行性的「Najash rionegrina」出現得更早。蛇類與滄龍有著相似的骨骼構造,彼此的四肢都已退化(甚至已消失),而且從解剖學的角度研究所得,二者之間的結構亦符合在支序分類學上的承接關係(雖然亦有一些特徵僅能顯示滄龍與巨蜥之間的聯繫)。不過,近年一些遺傳學的研究卻顯示蛇類與巨蜥其實並沒有如此親密的關聯,因此推斷蛇類與滄龍之間的關連亦同樣不見得緊密。雖然如此,但目前能將蛇類與滄龍作聯繫的生物證據,仍然比將蛇類與巨蜥作聯繫的證據為多。現在仍有一些從侏羅紀時代及前白堊紀時代所遺留下來的蛇類化石碎片正接受研究,由於目前的研究對於蛇類起源的推測尚未有確切的定論,因此任何新的研究進展都可能會推翻上述的任何一個假設。
現代蛇類主要分類的出現,大約始於古新世,亦即在恐龍滅絕及哺乳類動物面臨輻射適應階段時所發生。當時在北至斯堪的納維亞的北極圈,南至澳洲大陆及塔斯曼尼亞的所有地域裡,共出現了超過2,900個蛇種。[9]這些蛇類主要分佈在所有大陸上(南極洲除外)與及海洋之中,即使在高至海拔4,900米的亞洲喜馬拉雅山仍能找到牠們的蹤影[9][14]。不過,在某些島嶼上是完全找不到蛇類的,例如爱尔兰、冰岛及新西兰等[14]。
另外,世界上偶爾會發現一種奇特的生物現象:雙頭動物(甚至多頭)。當中比較容易出現雙頭情況的是爬蟲類動物及兩棲類動物,尤其是龜與蛇[15]。世界上曾經有數次報導指在個別地區發現雙頭蛇。2006年,美國水族館就曾經拍賣過一條罕見的雙頭蛇[16][17]。大部分雙頭蛇都只能存活一段很短的時間(約數個月),不過亦有一些報導指曾經有雙頭蛇以正常的蛇齡存沒,甚至能繁殖出正常的幼蛇來。一條黑色的雙頭錦蛇就據稱曾經活著20年之久[18]。
生物特徵
攝食及消化系統
所有蛇類都是肉食性的,牠們主要進食蜥蜴、小型哺乳類動物、鳥類、魚類、蛋類、其它蛇類、蝸牛及一些昆蟲[9][19][20]。由於蛇類並不能以牙齒將食物咬開及撕成細塊,因此牠們必須將食物整個吞下。蛇的體積直接影響牠們所選擇食物的大小,幼小的蛇類只吞食體型較小的獵物。例如幼蛇階段的蟒蛇一開始或者只能吞食蜥蜴或鼠類,長大後便能進食小鹿或者羚羊了。
蛇類的顎骨是眾多動物中最富韌力的。牠們的下顎相當有彈性,兩顎的接合位並不牢固,加上在頭骨周邊許多關節的輔助之下,令蛇類的兩顎能張開至足以把整頭獵物吞進口中,儘管獵物的直徑比蛇類本身還要大,蛇類的嘴巴仍能在不進行咀嚼的情況下將對方吞食下來[19]。例如食卵蛇就能把比自己頭型及體寬還要闊大的蛋吞下[20]。食卵蛇是沒有牙齒的,但在牠們的食道內側長有一些骨質的尖刺,能為牠們在體內把蛋殼捏碎,方便吞食[20]。
大部分蛇類都以進食小動物為主食,但某些品種的蛇卻是例外。例如王蛇和眼鏡王蛇就以進食其它蛇類為主;一種學名「Pareas iwasakii」的蛇以及新蛇科下的一種以蝸牛為食的鈍頭蛇,牠們的牙齒結構相當特別,口腔內右側的牙齒比左邊的多,這是因為蝸牛的外殼是呈順時針螺旋狀的,這種牙齒結構方便牠們把蝸牛咬開[21][22]。有些蛇類可以施展蘊含毒素的咬擊,這是為了在進食獵物之前先把牠們制服或殺死,方能放心地把對方吞進體內[19][23]。本身沒有分泌毒素的蛇類,則主要以身體擠壓獵物致死[19];更有些蛇類會把仍然未斷氣的獵物直接吞下[19][20]。
進食完畢後,蛇類會進入短暫的休眠狀態,靜待身體消化食物[24]。它們的消化過程是相當緊張的,尤其在剛吞下體型比較大的食物之後。某些偶爾才進食一次的蛇類,進食後其腸臟會慢慢地把食物保存並轉化成能量,這個過程一般都需要花費足足兩天才能正式完成。作為變溫動物,周邊的氣溫對於蛇類進行消化往往構成極大的影響,而讓蛇類進行消化的最佳溫度是攝氏30度。一種南美響尾蛇會在其消化過程中產生強烈的代謝能量,據紀錄所得這種能量會令牠的體溫上升至比周邊溫度更高出1.2度[25]。由於蛇類的消化過程是緊張的,所以當正處消化階段的蛇類受到騷擾的時候,牠們往往會先把食物吐出,以確保自己有足夠的能力逃離危險。只要沒有外在的騷擾,蛇類的消化效率會較高,因為蛇類的消化酶能溶解並吸收大部分食物;不過蛇類不可以把食物所附帶的毛髮及爪牙也消化掉,最後必須把這些殘餘物與尿酸一起排出體外。
蛇鱗及蛻皮習慣
蛇類的表皮是以蛇鱗所緊緊覆蓋的。很多人把蛇類的外在質感與蚯蚓的質感混淆,其實蛇皮的觸感是乾爽而平滑的。多數蛇類都以其鱗片來抓緊地面及進行移動。蛇鱗的質感可以是平滑的,也可以是骨質的或者粗糙而有微粒的。蛇類的眼瞼位置長有透明的眼膜,這片薄膜亦是蛇鱗的一種,英語稱為「Brille」。
蛇類有換皮的習性,一般被稱為「蛻皮」。在蛇類的蛻皮習慣中,蛇皮是一層疊一層的。[26]蛇鱗是緊密相連的,新舊的蛇皮緊緊相連,蛇類蛻皮的過程就像把襪子從腳掌褪出的情況一樣。[27]
蛻皮對於蛇類而言有很多功效。第一,最外層的受損舊皮得以替換;第二,換去蛇皮可以免於受蛆蟲等寄生蟲所侵害。在很多會換皮的生物(如昆蟲)例子,均證明蛻皮是令生物變得更巨大的手段,不過蛇類的蛻皮習慣是否有這種作用則尚在爭議當中。[27][28]

蛇類的蛻皮是每隔一段時間便會重覆進行的,而蛇類畢生都會一直進行蛻皮。在要蛻皮之前,蛇類會停止飲食並躲在某個安全的地方。蛻皮時蛇類的表皮會變得暗啞乾燥,牠們的眼睛會變得混濁或者呈現藍色。舊皮的內側面會產生水份,這令舊皮與新皮能夠得以分離。數天後,蛇眼重新變得清澈,而蛇皮亦已經脫下。舊皮是從嘴部開始被撐開的,蛇類透過不斷與外物(多是粗糙的地表或石頭)磨擦,使盡力氣從舊皮中穿出。很多時候舊的蛇皮都是從頭部至尾部向後方蛻去的,蛻下來的舊皮就像一隻褪出來的襪子一般,而舊皮底下的新皮則顯得更新、更大及更具光澤。[27][29]

一條年紀較大的蛇每年平均只進行一至兩次的蛻皮,而一條成長中的幼蛇一年可能會進行四次蛻皮。[29]只要蛻皮的過程是順利的話,被蛻下的舊蛇皮往往能留有清晰的蛇鱗形狀及分佈,從而能判斷出該蛇的品種[27]。蛇類蛻皮的主要目的是為了成長,但同時亦能去除身上的寄生物。這種定期替換新皮的概念被認為是象徵「治療」與「藥理」的標誌,因此象徵醫學的「蛇杖」標誌上,便有一條纏著木杖的蛇出現[30]。
蛇鱗在頭部、背部及腹部的各種分佈,是為蛇類分門別類的主要特徵之一。在西方,分佈在蛇體各部分的每種鱗片都有其專用的名字。對於一些較為進化的蛇類(例如新蛇),其腹部的鱗片列數是對應其脊椎數量的,這讓科學家不需要透過解剖就能掌握這些蛇類的脊椎段數。
點數蛇類鱗片還可以辨認蛇類的性別。在某個實驗中,學者曾把探針放進雌蛇的泄殖腔裡,紀錄雌蛇在動止間的泄殖腔位置。[24]實驗證明雌蛇泄殖腔跟雄蛇的半陰莖上的鱗片數字是有差別的(雄蛇的半陰莖比較長),因此點數鱗片的工作確實能判斷蛇類的性別。[24]
視力
蛇類的視力情況是令人注目的。一般而言,樹棲性的蛇類擁有較佳的視力,而居於洞穴內的蛇類視力則較差。蛇類最基本的視力是可以捕捉外界的動態,例如移動狀況,但未必能確切地予以掌握。某些蛇類像亞洲綠瘦蛇(瘦蛇屬的一個品種)一類,牠們是以雙目並用的模式來觀看的,牠們的雙眼能同時聚焦在同一點上。多數蛇類都只能前後移動晶狀體來進行聚焦,把影像模糊地投射在視網膜上;而其它脊椎動物的水晶體則是的放鬆,並能自由轉動的,因此蛇類的視力普遍而言是較差的。
嗅覺
蛇類以嗅覺追蹤獵物,可是牠們的嗅覺器官位於舌頭。蛇類的舌尖是呈叉狀的,牠們以舌頭收集空氣中的漂浮粒子,並將其傳遞至位於嘴部的犁鼻器(費洛蒙鼻嗅器)以測試氣味[31]。叉狀舌尖能同時刺激蛇類的味覺與嗅覺,為它們帶來追蹤味道的方向感[32]。一般蛇類都會讓舌頭長期保持運動狀態,一直測試著空氣中、土地上、水裡的各種粒子,以此找出獵物的所在位置與及探知身處的環境狀態[32]。
感應力

蛇類的身體長期接觸地面,牠們對於周邊的震動非常敏感,所以蛇類能夠透過身體感應空氣中及陸地上的震動,從而探測到附近有其它動物走近自己[32]。
另外,蝰蛇、蟒蛇及部分蚺蛇身體都有熱能感應器,位置在雙眼與鼻孔之間的一條深邃凹槽。另外,部分蟒蛇在上唇附近有一些小洞,讓牠們可以感應到外界放射的熱能[32]。熱能感應的能力讓蛇類能更準確地掌握獵物的位置,尤其當對方是恆溫動物的時候。
內臟
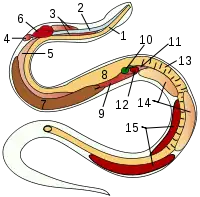 | 内部器官 | |||||||
|---|---|---|---|---|---|---|---|---|
| 1 | 食道 | 6 | 心脏 | 11 | 胰脏 | |||
| 2 | 气管 | 7 | 肝脏 | 12 | 脾脏 | |||
| 3 | 气管肺 | 8 | 胃 | 13 | 肠 | |||
| 4 | 原始左肺 | 9 | 气囊 | 14 | 睾丸 | |||
| 5 | 右肺 | 10 | 胆囊 | 15 | 肾 | |||
正如所有爬蟲動物一樣,蛇類也是變溫動物(即俗稱「冷血動物」)。蛇類的心臟包裹在一個囊中,稱為「心囊」,位於支氣管的分叉處,其心臟可以移動。另外,蛇類並沒有橫膈膜。這種調整是為了避免在吞服大型食物的過程中,其心臟可能會遭受損傷的潛伏危險。另外,蛇類的脾臟依附在膽囊及胰臟之間,功能是過濾血液。蛇類的胸腺位處心臟上方的脂肪組織層裡,負責製作血液中的免疫細胞。其心臟血管系統同樣非常獨特,蛇類的血液會從尾巴流經腎臟後,才再次回到心臟[33]。
蛇類的左肺在生物演變的過程中退化,一般顯得比較細小甚或並不存在。這是由於蛇類天生而來的管狀身體,基本要求體內器官的配合,才能適應其既瘦且長的獨特體型[33]。在大部分蛇類中,只有一個肺能發揮基本功能。牠們的肺部擁有血管化的前段及後段兩段脈管,這種結構並不適用於氣體交換的正常模式[33]。這個「囊狀的」肺主要用作應付水壓(流體靜力學),一些在水中活動的蛇類倚靠肺部來調整身體的浮力。而對於在陸地上活動的蛇類而言,這個肺部的實際功能仍有未能解釋的地方[33]。蛇類有很多器官都成對,例如腎臟及生殖器官等,都在身體內不分左右,只以前後分序地存在著[33]。另外,蛇類體內並沒有膀胱及淋巴結。
運動方式
蛇類生物在進行陸地移動時的主要方式是「蛇行」。蛇類生物沒有足部,而是利用腹鱗及肌肉的縮放產生移動力。其移動方式主要有四種,分別是蜿蜒式、側行式、直蠕式和風琴式。至於在水中生活的海蛇,一般尾部都會像魚類尾巴般扁平,當海蛇在水中進行蜿蜒式蛇行動態時,尾部便可如鰭、蹼般充當划水的推進器。
繁殖

雖然蛇類的繁殖模式種類豐富,但所有蛇類都是體內受精的。雄性蛇類擁有一對呈叉狀的半陰莖,平時反進體內收藏在其尾巴之中[34]。雄蛇的半陰莖通常長著凹槽、倒勾或尖刺,目的是要在交配的時候能以性器官抓緊雌蛇的泄殖腔壁[34]。
多數蛇類都是卵生的,而且雌蛇都會在誕下蛇卵不久後便遺棄掉它們。可是,亦有部分蛇類會認真地築起巢穴並守在蛇卵附近孵育它們,如眼镜王蛇[34]。蟒蛇則會在誕下蛇卵後以身體捲著它們,直至幼蛇孵化出來[35]。除了偶爾為要增加體溫以助孵蛋因而需要曬太陽及喝水之外,雌性的蟒蛇都不會離開蛇卵[35]。蛇為变温動物,體溫不足以孵卵,所以雌蛇會以反覆肌肉收縮的方式摩擦生熱來孵蛋。
有些蛇類是卵胎生的,蛇卵會保存在母體之內,準備孵育的工作[36][37]。最近,更有研究證明有一部分的蛇類是完全屬於胎生的,例如紅尾蚺及森蚺,牠們把胎兒養育於胎盤中,並為幼胎提供營養,這種情況在爬蟲類動物中,甚至在真獸下綱以外的其它物種中也是相當反常的現象[36][37]。另外,蛇類的生產多數都在溫度較低的環境下進行[34][37]。
蛇毒

眼镜蛇科的蛇、蝰蛇科的蛇及部分游蛇科的蛇類等都以毒素來制服並殺死獵物。這些毒素是蛇類變種的唾液,從附在牙齒後的毒囊所分泌並傳至牙齒之內[38]。毒牙位於口腔前部的蛇類的牙齒或者是空心的,或者具有溝槽,這令毒液注射的過程更加有效率;而某些蛇類(如樹蛇)只有位於後方的牙齒內才藏有傳輸毒素的管道,被稱為「後齒」(Rear-fanged)的毒蛇。蛇毒對於大部分獵物都有特效,除了方便獵食之外,蛇毒亦是毒蛇類的重要自衛手段[38]。蛇類的毒素就像一般的唾液,充當一種前消化劑(有助消化的物質),它能分解食物為可溶複合物質,轉化為適合消化的成分;即使是無毒的蛇類,也會令食物的組織受到破壞,因此幫助消化的體液是必須的[38]。不過據研究,一些恆常作為毒蛇獵物的鳥類、哺乳類動物及其它蛇類,可能會發展出抗毒甚至完全免疫的體質[38]。
分辨有無毒性的方法,“通常觀察其頭部,若是三角形狀,該種類便有毒,反之若是圓形便無毒”其实是分辨是否為蝰蛇科蛇类的方法,但还有一大类眼镜蛇科蛇类并非“三角头”,因此并不全面,不过在中国内陆,多数毒蛇都属蝰蛇科,此方法经常有效。毒蛇包括三個主要的品種(下文將會介紹),不過牠們之間並未組成正式的毒類組別。在某些場合的英語中,以「poisonous」一詞形容蛇毒其實是不正確的,因為「poison」所指的是以「吸進」及「攝取」方式傳達的毒素,而蛇的毒素是「注射」式的,因此應該用「venom」一詞較為恰當[39]。不過,亦有兩種蛇的毒素來源是較為特殊的:頸槽蛇會在吞食蟾蜍時阻隔其毒素於頸部,並從處於其頸部的分泌腺分泌出那些毒素,藉以作為保衛自己的武器;在俄勒岡州有少量束帶蛇(新蛇總科的一個品種)會在肝臟保存著因進食蠑螈所留下的毒素,並以這些毒素作為自衛手段,藉以與當地的掠食者(如狐狸、烏鴉等)抗衡[40]。有一種被分類為射毒眼鏡蛇(如紅射毒眼鏡蛇)的蛇類,能在一定距離內透過毒牙向敵人噴射出毒液,這些毒液的傷害性比較弱,但如果雙眼被毒液濺到,卻足以損害生物的視力,其毒性足以造成失明。
蛇毒一般是以蛋白質為主的複合物質,平常貯存在頭部後方的毒素腺中[40]。所有毒蛇體內的毒素腺都會透過體內的管道,把毒素傳送到上顎的空心牙齒中[38][39]。毒素中的蛋白質大都是神經毒素、肌肉毒素及細胞毒素等多種毒素的混合物質,這些毒素會直接攻擊生物的神經系統及肌肉系統,亦可能導致呼吸系統障礙、肌能麻痺,最終令生物死亡[39]。幾乎所有蛇毒都蘊含「玻璃酸酶」,這是一種會令毒素迅速擴散的酶[38]。當被蛇咬到時,不可快速奔跑或劇烈運動,應火速就醫,劇烈運動只會加速駛毒液流入心臟。
使用肌肉毒素的毒蛇主要以尖銳的前齒分泌蛇毒,並注入獵物的體內,令其失去反抗能力[39]。有些毒蛇則使用神經毒素,例如貓眼蛇,牠們的毒素位於後排的牙齒,而且這些牙齒是向後方(向體內)彎曲的[41],這類毒蛇難以運用牠本身的毒素,而對於科學家而言亦較難搾取牠們的毒液[39]。眼鏡蛇及環蛇是屬於以前排牙齒分泌毒素的種類,不過牠們的空心牙齒並不是向前直伸的,因此牠們不能像蝰蛇般以牙「刺」或「戮」向對手,只能以「咬」的方式進行攻擊[42]。
最近有一個新的看法,認為所有蛇類都可能是有毒的,一些蛇類只是因為能分泌的毒素太弱或沒有牙齒的原因,才被普遍認為是沒有殺傷力的蛇種。[43]蛇類的毒素也許是承繼自蜥蜴類的先祖,現今存在的赫拉毒蜥及墨西哥毒蜥便是帶有毒素的蜥蜴類代表。毒蛇與毒蜥都是蜥蜴類動物的直接毒素繼承者。
毒蛇主要分成兩個主要種類:
另外,還有第三種有著「後齒」結構的蛇類亦是毒蛇:
一般會將蛇毒素分為兩類:出血性與神經性。臺灣的毒蛇中,雨傘節是「神經性」,百步蛇是「出血性」。
與人類的互動
蛇咬
蛇類一般不會視人類為獵物,多數蛇類都不會主動攻擊人類,除非牠受到驚嚇或傷害,才會發動攻勢,否則一般而言蛇類都會避免與人類發生接觸。除了一些巨大的蛇類(如蟒蛇、蚺蛇)外,大多數無毒的蛇類都不會對人類帶來生命上的威脅。無毒蛇類的咬擊通常沒有太大殺傷力,因為牠們的牙齒主要是用作抓住多於撕開獵物,因此普遍不會造成較深的傷口。雖然被蛇所咬的皮膚及肌肉組織會有所損害,但相比於毒蛇的咬傷而言,無毒蛇類的咬傷較無重大威脅。[38]
被蛇所咬而致命的紀錄實際上並不算多,然而少數受毒蛇咬傷且缺乏急救的情況下可能導致肢體切除手術以避免致命。全球約有725種毒蛇,當中只有250種毒蛇能以一口咬擊立即奪人性命。雖然澳洲是全球最多毒蛇聚居的地方,但平均一年只有一遭咬擊是由有毒蛇類所導致的;在印度,一年約有250,000多宗蛇咬案件,其中有50,000宗導致死亡,多數案例是由於缺乏適當或及時的急救措施[44]。
醫治蛇咬的方法種類繁多,最典型及常用的方法是運用由蛇毒本身製煉的抗毒血清。一些抗毒血清是專為個別及少數近類蛇種而用的(例如在美國當地,基本上除了珊瑚蛇外,所有毒蛇都屬於蝮亞科),而另一種能治療多種蛇毒的血清是「蛇毒血清」(antivenin polyvalent)。製作蛇毒血清時,研究者會把多種毒蛇的毒素同時注入一匹馬的體內,劑量由輕至重逐步提升,直至馬匹能對毒素免疫為止;接著便從馬匹身上抽取血液,再以淨水加以稀釋,並冷凍貯存,最後成功製造血清。可是亦因為這個原因,一些對馬匹過敏的人便不能使用這些血清。在印度、南非及澳洲等地,當地人亦針對較為危險的蛇類(如曼巴蛇、太攀蛇及眼鏡蛇),以同樣手法製作特種抗毒血清。
弄蛇術

在世界上的某些地方,尤其是印度,弄蛇術是相當著名的街頭表演。在這些表演中,弄蛇者會帶著一個放有蛇類的籃子,透過吹奏樂器(笛)而令蛇類作出各種特殊的反應[45]。其實蛇類是沒有外耳的,儘管牠們長有內耳,但牠們並不會受到音乐的影響[45]。
有研究者指出,許多弄蛇者實際上是以雙手的姿勢向蛇類發出訊號,蛇類其實是根據笛子的左右擺動與及弄蛇者跺腳的震動才會作出多種扭動身體的動作。而弄蛇者的這些技巧多數不易被在場的觀眾看出,因為他們大部分都是厲害的障眼法高手。弄蛇者很少會直接用手捉著他們的蛇,另外他們的蛇亦大多數是無毒的或是被剝下牙齒的眼鏡蛇。有部分弄蛇者會表演眼鏡蛇與獴隻相鬥的環節,不過這種環節並不普遍,因為不管是蛇類還是獴隻都很容易會在相鬥的過程中受傷,甚至死亡。現在印度政府為了保護野生蛇類,決定停止國內的街頭弄蛇表演業。事實上,在一些印度地區內,弄蛇業是牴觸法律的。[45]
捕蛇業
在印度的安得拉邦及泰米爾納德邦附近的部落「伊魯拉斯族」(Irulas)有世代相傳的獵蛇者及捕蛇習俗。他們在乾燥炎熱的森林裡,經常鍛鍊捕蛇的技能,並有著關於當地蛇類的豐富知識,憑一根簡單的棍棒便可以捕蛇。在早期階段,伊魯拉斯族人能定時捕捉過千條的蛇類,並以蛇皮為材料進行多種工業活動。1972年,印度當地實施「野生動物保護行動」,禁止蛇皮工業,令伊魯拉斯族人不能再倚靠這種工業維生;於是伊魯拉斯族中的捕蛇者便組織成聯盟,專門捕捉毒蛇並把牠們的毒素去除,再將牠們放生。他們所收集的毒液便成為了製作抗毒血清的重要材料。[46]伊魯拉斯族人所捕捉的蛇,還可作食用及捕鼠之用。
儘管在印度當地已有不少弄蛇者的存在,但亦有很多專業的捕蛇者同時作業。現在透過爬蟲學的研究得出結論,就是捕捉蛇類最理想的工具是一根末端呈「V」字型的長棍,不過亦有一些人物如比爾·哈斯特、奧斯甸·史提芬斯與及傑夫·可雲等人主張徒手捕蛇。
蛇類的用途

在一些飲食文化中,蛇肉是可以被食用的,甚至被認為是一種美食。徐珂《清稗类钞》补载:“粤人嗜食蛇,谓不论而蛇皆可佐餐,以之缕丝而作羹,不知者以为江瑶柱也,盖其味颇似之,售蛇者以三蛇为一付,易银币15元,调羹一,须六蛇,需30元之价矣……其以蛇与猫同食也,谓之曰龙虎菜;以蛇与鸡同食,谓之曰龙凤菜。”蛇羹在廣東菜中是在秋天時進補抵寒的有益食物。在中医学的角度而言,進食蛇類有心臟保暖的功能。據記載,西方文化卻只有在非常飢餓的情況下才會進食蛇類[47]。然而在美國的中西部,煮熟的響尾蛇肉卻是普遍的食材。在大部分亞洲國家諸如中國、泰國、印尼、越南及柬埔寨,都有飲用蛇血的習慣,據稱這能加強雄性的性能力[48]。如果做得到的話,當地人更會在仍然存活的眼鏡蛇身上取血,並混以酒水作調味。另外,亞洲亦有以蛇浸酒的做法,通常會把整個蛇體放進酒埕裡,據說這樣能加強酒的濃度(價值亦因此更昂貴)。在日本,有一類名叫「波布」的蛇類,主要分佈在沖繩一帶,當地人會把牠們製作成「波布清酒」[49]。
在中國、拉丁美洲、中南半島都有吃蛇的習慣,並認為吃蛇有滋陰補陽、清血美膚效果;廣東菜譜有深秋食用「菊花五蛇羹」及「太史五蛇羹」[50](秋天蛇即將冬眠,體質最肥美)、「龍虎鬥」(蛇肉貓肉同燉)或「龍虎鳳」(蛇肉、貓肉及雞肉)。[51]日本人把蛇製成蛇粉、酒來補身。另外,蛇膽亦號稱具有藥用價值,可是味道甚苦。[52]生食蛇胆有很大风险感染裂头蚴。相反,在猶太教中的飲食規則,蛇被列為不淨動物而不建議食用。
另外,在世界很多地區都有人飼養蛇類。在西方國家,球蟒、王蛇屬及錦蛇屬一類性情較溫和的蛇類是受歡迎的寵物品種。為了供應作寵物用途的蛇類,一些人工飼養及繁殖爬蟲類及兩棲類動物的行業便應運而生,被飼養的蛇類比起野生蛇類更適合當寵物。另外,蛇皮亦具備裝飾用途,許多盛產蛇類的地方都會抽取蛇皮製作成各種皮具用品,如錢包、手提包等。不少胡琴(如二胡)以及日本傳統弦樂器三味線的琴身,亦是以蛇皮製作的。[53]
相關文化
中國大陆



「蛇」字作聲旁的「它」字,本來是蛇的象形文字,而其部首「」字的讀音,也與蛇的別稱「虺」一樣。[54]在東亞文化圈的十二生肖中,蛇排在第六位,對應的地支巳字,也是蛇的象形文字。[55]蛇類在中國的形象頗為複雜,既具神格,亦具妖格。蛇有特殊的原始地位,人類始祖伏羲與女媧是半人半蛇的形態。[56]蛇亦被稱為「小龍」,有時被視為是龍的化身,王充在《論衡》說:「龍或時似蛇,蛇或時似龍。」又說:「龍鱗有文,與蛇為神。」鄭玄注《尚書大傳》亦指:「蛇,龍之類也,或曰:龍無角者曰蛇。」《後漢書·襄楷傳》有「夫龍能變化,蛇亦有神,皆不當死」之語,都表示古代的蛇亦有龍像,故此具備神格。中國道教大神玄天上帝腳踩龜將、蛇將各一,四方神獸之一的「玄武」也就是龜蛇合一的形象[57][58];中國神話人物夸父身上有蛇為裝飾,《山海經》中更常有上古傳說人神或生物以蛇類為飾物的記載,在書中經常出現「珥蛇」[59]、「操蛇」或「踐蛇」的描述;中國一眾神獸之中,亦有名為「螣蛇」的蛇神,又有能興雲布雨的黑蜧;西漢開國君主劉邦曾經「斬白蛇起義」,白蛇被指為「白帝子」;[60]《晉書》記載晉朝開國名將杜預曾因酒醉而化成大蛇,因而被傳說為大蛇轉世[61](宋代時人魚周詢同樣有類似傳說[62])。在晉代以前,蛇有時會被理解為吉祥的象徵。《搜神記》記載春秋時代隨國君主隨侯(或作隋侯)曾因救治一條蛇,而獲靈蛇銜珠相贈,以作回報,「靈蛇珠」或「隨侯珠」從此成為中國古代珍貴寶物的代表之一,與和氏璧齊名。[63]西晉文人傅玄曾作《靈蛇銘》:「嘉茲靈蛇,斷而能繼。飛不須翼,行不假足。上騰霄霧,下遊山岳。進此明珠,預身龍族。」銘中提及螣蛇形象、靈蛇贈珠之事,「行不假足」的特性,與及「斷而能續」的技能,是一首將蛇的形象神化的文學作品。[64]民間百姓供奉的「五大仙」(又稱「五大家」或「五顯財神」)中,蛇仙被稱為「柳仙」,形象介乎妖仙之間。道教文獻《道法會元》裡,有靈將名叫馬元帥(即馬靈官或馬天君),又有副將馬充,祂們都有著「紫庭追蟲下瘵凝神化氣轟天素練白蛇大元帥」、「都天捲祟白蛇大將」、「轟雷霹靂練素白蛇大將」、「上清正一火犀大仙南極靈官金睛馘魔威烈元帥」等仙銜,亦有所謂「召白蛇咒」、「白蛇符」等咒文,是道教中以白蛇為原型的著名神靈。
漢民族有蛇圖騰崇拜,如女媧為半人半蛇之形象,亦有女媧造人之說。閩越族亦有如此,漢代《說文解字》云:「閩,東南越,蛇種」,可知閩地之蛇圖騰崇拜,至今福建各地尚有崇蛇習俗,如南平市樟湖鎮有蛇王廟、福州市長樂區潭頭鎮岱西有蟒天神王廟、長樂區閩江口厚福英武廟英烈王等。
另一方面,蛇在中國的妖怪形象頗多,一些誌異作品中蛇精修練得道的故事,出現頻率僅次於狐狸精。中國著名民間傳說《白蛇傳》中所提及的白蛇與青蛇,是具備人格的蛇妖的著名代表。志怪小說《聊齋誌異》中亦有很多以蛇為主題的妖怪故事,如〈豢蛇〉、〈青城婦〉、〈蛇人〉、〈斫蟒〉、〈海公子〉等篇。清代煙霞主人所寫的小說《躋雲樓》中有一首《鐵鞭蛇賦》,更形象化地將妖蛇描述出來。[65]蛇自古就有著多種類型的負面或妖物形態,戰國時代楚國詩人屈原的作品《天問》中,曾提及「一蛇吞象,厥大何如」,這典故其後發展成《山海經·海內南經》中著名的「巴蛇吞象」故事,而蛇亦被人賦予貪婪、食量驚人的印象。[66]而同在《山海經》中,尚有許多以蛇為原型的妖怪,例如有長蛇、化蛇、枳首蛇、肥遺、相柳等。晉代郭璞有許多文學作品均有賦蛇,當中的《長蛇贊》[67]、《巴蛇贊》[68]、《蟒蛇贊》[69]、《枳首蛇贊》[70]等大都刻畫形象較負面的怪蛇。

五行概念中,蛇屬巳,有火相。[71][72]二十八宿中有翼火蛇。在民間,俗稱「五毒」的生物當中包括毒蛇(其餘為蠍子、蟾蜍、蜈蚣和壁虎,後改以蜘蛛取代壁虎);[73]民間武術的「五形拳」中有「蛇拳」。[74][75]在廣東語言文化裡,蛇有著「懶惰」的表現,因此指人偷懶會稱其「蛇王」;另外,在撲克牌遊戲大老二中,五張數字順連的牌型亦稱為「蛇」或「一條蛇」。
中國古代軍事學上,有所謂「一字長蛇陣」。《孫子兵法·九地篇》載:「故善用兵者,譬如率然;率然者,常山之蛇也,擊其首,則尾至,擊其尾,則首至,擊其中,則首尾俱至。」是一種首尾相顧的長線形戰略陣式。《管子·兵法》篇中提到「九章」的概念,以不同的旗章提示軍隊不同的行軍模式,當中「舉蛇章,則行澤」表示蛇形旗章是行軍於河澤之意。[76]另外,唐朝常州刺史獨孤及所著的《八陣圖記》,記載所謂「八陣圖」中的八陣,當中包括「蛇蟠陣」。小說《封神演義》及《薛家將》中提及所謂「十陣」,當中亦有「一字長蛇陣」。
日本
日本最著名的蛇神是神話中的妖蛇八岐大蛇。七福神中的辯才天也有蛇的特徵。[77]別名「三輪明神」的大物主神,兼具蛇神、水神與雷神的神格。茨城縣當地亦有對蛇神夜刀神的崇拜。日本部分地區有尊奉白蛇的信仰習俗,如岩國白蛇。另外,也有很多關於「蛇女」傳說的故事(如著名的蛇女房)。[78]妖物方面,日本有蛇骨婆、野槌、手負蛇等著名的蛇型妖怪。陰陽家安倍晴明所撰《占事略決》中有十二天將的概念,當中有「騰蛇」,五行中屬火神凶將(與中國天干五行屬性相若),家在巳,主驚恐怖畏。日本戰國時代軍事家武田信玄所提及的十陣,亦有「長蛇陣」。
臺灣原住民
排灣族:對他們而言,「百步蛇」是祖靈的象徵,稱其為tasalad(夥伴)、kavulungan(祖先)等,據說石板屋建築就是受到蛇鱗的啟發。 魯凱族:在他們的傳統故事中,也有類似於祖靈的象徵,皆是由蛇開啟他們的生命。 布農族:對她們而言,百步蛇被視為朋友(Kavuaz)例如在「百步蛇的復仇」這篇故事中,就有和蛇(動物)講信用的例子。 邵族:在他們的故事當中,描述的是蟒蛇,在「蟒蛇泅水」中就有提到蟒蛇的造形給予他們製作獨木舟的靈感,因此「蟒甲」即為原住民族語中的「獨木舟」之意。
印度
印度是以崇拜蛇而著名的國度之一,其創世神話「乳海攪拌」中眾神就是以蛇神婆蘇吉為攪拌乳海的工具。印度一帶著名的蛇則以印度眼鏡蛇為主,主神濕婆頸上常纏著一條護法的眼鏡蛇,大神毗濕奴就與千蛇之王舍沙有著密切的關係;[79]另外,印度教及佛教中亦有不少以蛇為原型的神祇,例如那伽、摩納娑、摩睺羅伽,佛教神明軍荼利明王身上有蛇為裝飾。
世界各地文化的蛇

蛇在世界各地的文化中都飾演著很特殊的角色。牠既有邪惡的一面,卻又具備醫學意義甚至神格,在某些地區受到民族性的崇拜(例如在位於加勒比海的法國海外大區馬提尼克是雙線細盲蛇的標本產地)。在英語中,除使用「Snake」作為蛇的名稱外,亦有較為文學化的稱謂「Serpent」,因此許多西方文化都以「Serpent」來稱呼神話及傳說中的各種蛇類。而在西方諸國的不少神話傳說中,都有「大海蛇」這種形象的存在,如舊約聖經裡的利維坦,[80]與及北歐神話裡的耶夢加得,[81]同樣象徵邪惡的力量。在西歐為主的基督教文化圈裡,大部分情況下蛇被視為邪惡的象徵,因為蛇曾誘惑夏娃吃禁果;而亞當的第一任妻子吸血鬼莉莉斯亦有蛇的意味。[82]。在舊約出埃及記中,摩西的權杖依照耶和華之意變成蛇出現在法老面前(出埃及記7:10)。但圣经中亦有视蛇为救赎化生的地方,如《民数记21:8》:以色列民旅行在旷野时,耶和华用一条铜蛇治愈大毒蛇的毒液,它在新约圣经中仍然预示着十字架上的基督(《约翰福音3:14》)。 世界上還有不少具代表性的蛇形圖騰,如自繞一圈叼著自己尾巴的「銜尾蛇」圖案,是現代數學概念無窮的原型。[83]另外,基督教普遍認為「銜尾蛇」是整個物質世界的邊境與限界,其「自我消減」的特性也象徵著追隨《傳道書》中的傳道者足跡,在轉瞬即逝的世界裡,一個短暫的有限的存在。[83]卻斯特頓在《永存者(The Everlasting Man)》中便以銜尾蛇作為「循環定律」的標誌,是泛神論與神秘主義中「自我毀滅」的代表。[83]銜尾蛇在匈牙利及羅馬尼亞等東歐國家的獨神論派系教堂中,是一種重要的徽紋。[83]東歐基督教有一個俗稱拜蛇教的支派,信奉的就是銜尾蛇,並稱其為「聖蛇」。[83]而在諾斯底主義中,銜尾蛇則象徵「無限」與及「世界之魂」。[83]《哈利波特》一書中的學院史萊哲林的象徵物也是蛇。

世界衛生組織旗幟上的圖徽中亦有一條蛇,源自於古希臘醫神阿斯克勒庇俄斯的權杖,因為蛇類蛻皮重生的特性被視為「醫學智慧」象徵的體現,因此「蛇杖」象徵「醫學」;而阿斯克勒庇俄斯的女兒許癸厄亞則是希臘的健康女神,手持一隻藥碗,碗上同樣纏著一條長蛇,象徵「藥學」。[84]
希腊神话中,有很多關於蛇的神祇及與蛇相關的故事。在神話世界中本來擁有頗高神格的厄客德娜,是一頭上半身是妖艷女性,下半身卻是大蛇的妖怪;宙斯的其中一個情婦拉彌亞也是一頭半人半蛇的妖怪。[85]大力士海格力斯的十二大偉蹟中,其中一項是殺掉沼澤裡的九頭蛇;[85]著名的蛇髮女妖戈耳工及美杜莎等,都是以蛇為形象的妖怪。[85]在神話中,蛇髮女妖美杜莎的雙眼能令看到她的生物石化,最後她被珀耳修斯取下首級獻給女神雅典娜,雅典娜將其嵌在神盾埃癸斯中央,使神盾保持令對手石化的能力。這個傳說常被後世創作所採用(如奇幻小說《哈利波特》系列及《波西傑克森—神火之賊》、《惡魔城》系列、電影《超世紀封神榜》、《魔法戰隊魔法連者》的冥府神戈耳工、《ONE PIECE》的九蛇女帝波雅·漢考克、《FAIRY TAIL》的艾芭葛琳、《聖鬥士星矢》的英仙座、《Fate/stay night》中的英靈Rider、《飛輪少年》的美作涼等)。此外,一些著名的傳說生物如翼蜥、飛龍等都以蛇為形象原型。
值得一提的是,世界各地神話傳說中的大海蛇,不管是北歐神話的耶夢加得、歐洲文化中的利維坦(有時形象是大蛇)、海德拉,還是日本的八岐大蛇,都被視為當地的水神,牠們發怒時會引發氾濫、海嘯等災難,所以是各地人民又敬又畏的對象。
参考文献
- 《南美出土13米長巨型蟒蛇化石 (页面存档备份,存于)》,BBC中文網,2009年2月4日.
- S. Blair Hedges(August 4, 2021).“At the lower size limit in snakes: two new species of threadsnakes(Squamata: Leptotyphlopidae: Leptotyphlops)from the Lesser Antilles (页面存档备份,存于)”(PDF).Zootaxa,1841:1–30.於2008年8月4日查閱
- 康加(1991年),頁23。
- . ITIS. 2007 [12 December, 2007].
- . ITIS. 2007 [13 December, 2007].
- . ITIS. 2007 [13 December, 2007].
- . ITIS. 2007 [13 December, 2007].
- 美荷頓斯(1987年),頁11。
- 珊徹茲, 亞歷贊德羅, , 珊徹茲老爸的西印度自然歷史網站, [2007年11月26日], (原始内容存档于2016年3月4日)
- 麥道衛, 森姆爾, , 生物進化論, 1972年, 第6冊: 頁191至273
- 亞匹斯達古爾亞, 思巴斯地恩; 赫臣‧渣荷. . 自然. 2006年4月, 第440冊 (第7087期): 頁1037至1040 [2008-05-04]. doi:10.1038/nature04413. (原始内容存档于2007-12-18).
- 美荷頓斯, 羅拔, , 沙勞越博物館期刊, 1961年, 第10冊: 頁320至322
- . UNEP WCMC資料庫. 美國華盛頓: 美國先進科學聯盟. [2007年11月29日]. (原始内容存档于2007年12月25日).
- 康加(1991年),頁143。
- 亞連拿, 絲域連拉. . Pravda.ru. 2003年11月29日 [2006年12月31日]. (原始内容存档于2007年1月25日).
- . [2008-05-04]. (原始内容存档于2008-05-28).
- . BBC新聞. [2006年3月10日]. (原始内容存档于2020年5月17日).
- . [2008-05-04]. (原始内容存档于2012-11-02).
- 巴布拉(1979年),頁581。
- 美荷頓斯(1987年),頁81。
- M‧何索、T‧高廣及M‧堀《Right-handed snakes: convergent evolution of asymmetry for functional specialization》頁169至172,2007年。
- 美荷頓斯(1987年),頁184。
- 費利堡(1984年),頁125至127。
- 羅臣福(1989年),頁11。
- 格連‧J‧鐵達莎爾、威廉‧K‧米爾森、奧卡斯‧S‧安倍、西蒙爾‧P‧布列圖、丹尼斯‧V‧安德拉迪. . 生物實驗期刊 (生物學家出版社). 2004年, (第207期): 頁579至585 [2008-05-04]. (原始内容存档于2005-11-30).
- 史密夫、馬爾哥姆‧A:《Fauna of British India:Vol I - Loricata and Testudines》,頁30。
- 蛇是否黏呼呼的? (页面存档备份,存于)2006年8月14日
- . [2008-05-04]. (原始内容存档于2016-01-16).
- . [2008-05-04]. (原始内容存档于2007-11-25).
- 威爾科斯, 羅拔‧A; 域咸, 艾瑪‧M. . 醫學年報. 2003年4月15日 [2008年5月4日]. (原始内容存档于2020年6月24日).
- 大英百科:犁鼻器
- 康加(1991年),頁180。
- 美達, 德格拉斯, , 爬蟲類, 1995年6月, 第三冊 (第二期): 頁84至93
- 卡普拉(1989年),頁117。
- 康加(1991年),頁186。
- 卡普拉(1989年),頁118。
- 康加(1991年),頁182。
- 美荷頓斯(1987年),頁243。
- 費利堡(1984年),頁125。
- 費利堡(1984年),頁123。
- 費利堡(1984年),頁126。
- 美荷頓斯(1987年),頁242。
- 富萊, 拜仁‧G; 維度爾, 尼古拉斯; 諾文, 珍尼特‧A; 溫克, 費列克‧J; 史基保, 荷爾格; 拉姆傑恩, 拉恩; 科魯普, 辛查耶, , 自然, 2006年, 439: 584–588, doi:10.1038/nature04328
- 辛哈, 科堤雅, , 印度時代雜誌, 2006年7月25日 [2008年5月4日], (原始内容存档于2011年8月21日)
- 巴古拉, 帕拉華. . 国家地理杂志. 2002年4月23日 [2007年11月26日]. (原始内容存档于2007年12月18日).
- 維他可‧羅姆魯斯及卡普頓‧亞斯荷格:《Snakes of India: The Field Guide》頁11至13,2004年。
- 艾雲‧F‧R:《Snakes as food for man. British Journal of Herpetology》頁183至189,1954年
- 費連, 奧潔尼. . 傳說旅程. 2002年4月23日 [2007年11月26日]. (原始内容存档于2007年11月17日).
- 亞倫, 大衛. . Stars & Stripes. 2001年7月22日 [2007年11月26日]. (原始内容存档于2007年11月28日).
- . [2008-05-04]. (原始内容存档于2014-02-25).
- . [2015-10-27]. (原始内容存档于2008-10-19).
- (PDF). [2022-07-20]. (原始内容 (PDF)存档于2017-01-09).
- . [2008-05-04]. (原始内容存档于2008-12-08).
- . [2011-11-11]. (原始内容存档于2012-11-28).
- 《說文解字·巳部》:「巳:巳也。四月,陽气巳出,陰气巳藏,萬物見,成文章,故巳為蛇,象形。」
- 《後漢書·周燮傳》:「伏羲牛首,女媧蛇軀,臯繇鳥喙,孔子牛唇,是聖賢異貌也。」
- . [2008-05-04]. (原始内容存档于2008-05-13).
- . [2008-05-11]. (原始内容存档于2008-05-16).
- 晉代郭璞注《山海經》指:「珥,以蛇貫耳也,音釣餌之餌。」
- . [2012-07-09]. (原始内容存档于2012-07-09).
- 《晉書·杜預傳》:「(杜)預初在荊州,因宴集,醉臥齋中。外人聞嘔吐聲,竊窺於戶,止見一大蛇垂頭而吐。聞者異之。」
- 《宋史》列傳第六十一。
- 見《搜神記》及《藝文類聚·鱗介部》。
- 嚴可均輯:全晉文 (页面存档备份,存于)
- 原文:「維毒出之滋長,實變幻其莫測。苟考辨之未詳,每躬逢而受災。爾乃品居蛇類,名號鐵鞭。眠於夏日,旺在秋天。啖腦汁之一飽,恒掩臥乎三年。形雖蜿蜒,質同金鋼。掉尾相擊,所經必傷。伏行旅之邸舍。入佳人之閨房,時潛藏於林底,亦隱蟠夫屋樑。至若性忌燈光,喜托暗室。搏閃醉後,噬乘睡餘。不畏刀劍,專怕熏炙。雖產育乎此地,實土人所未悉。」
- 原文:「巴蛇食象,三歲而出其骨,君子服之,無心腹之疾。」意思是有一種巴蛇,能吞下整頭大象,經歷三年後才將象骨吐出;如果君子食用此蛇,可醫治一切心腹之疾。
- 《長蛇贊》:「長蛇百尋,厥鬣如彘。飛群走類,靡不吞噬。極物之惡,盡毒之厲。」見《藝文類聚·鱗介部》。
- 《巴蛇贊》:「象實巨獸,有蛇吞之。越出其骨,三年為期。厥大何如,屈生是疑。」見《藝文類聚·鱗介部》。
- 《蟒蛇贊》:「蠢蠢萬生,咸以類長。惟蛇之君,是謂巨蟒。小則數尋,大或百丈。」見《藝文類聚·鱗介部》。
- 《枳首蛇贊》:「夔稱一足,蛇則二首。少不知無,多不覺有。雖資天然,無異駢拇。」見《藝文類聚·鱗介部》。
- 王充《論衡·物勢》:「巳、火也,其禽虵(蛇)也。」又曰:「水勝火,故豕食虵。」又曰:「巳、虵也,申、猴也。火勝金,虵何不食獼猴?」
- 《西遊記》第六十六回「諸神遭毒手,彌勒縛妖魔」中有「龜蛇生水火,妖怪動刀兵」句。
- . [2008-05-11]. (原始内容存档于2008-06-26).
- . [2004-08-16]. (原始内容存档于2004-08-16).
- . [2008-05-04]. (原始内容存档于2001-09-23).
- 《管子·兵法》:「九章:一曰舉日章,則晝行。二曰舉月章,則夜行。三曰舉龍章,則行水。四曰舉虎章,則行林。五曰舉鳥章,則行陂。六曰舉蛇章,則行澤。七曰舉鵲章,則行陸。八曰舉狼章,則行山。九曰舉韟章,則載食而駕。九章既定,而動靜不過。」
- . [2011-11-06]. (原始内容存档于2012-12-24).
- Shinowa Chiwaki:日本幻奇文學第45章:蛇女傳說
- . [2008-05-11]. (原始内容存档于2022-04-06).
- . [2008-05-04]. (原始内容存档于2018-09-01).
- . [2008-05-04]. (原始内容存档于2013-12-26).
- . [2008-05-04]. (原始内容存档于2008-05-18).
- 銜尾蛇介紹
- . [2008-05-04]. (原始内容存档于2020-06-24).
- . [2008-05-04]. (原始内容存档于2008-04-30).
- . [2008-05-11]. (原始内容存档于2008-06-26).
- . [2008-05-04]. (原始内容存档于2009-04-18).
- . [2008-05-11]. (原始内容存档于2020-06-15).
- . [2008-05-04]. (原始内容存档于2021-05-06).
延伸閱讀
- Behler, John L.; King, F. Wayne. . New York: Alfred A. Knopf. 1979: 581. ISBN 0-394-50824-6.
- Bullfinch, Thomas. . London: Chancellor Press. 2000: 679 [2012-09-01]. ISBN 0-7537-0381-5. (原始内容存档于2009-02-09).
- Capula, Massimo; Behler. . New York: Simon & Schuster. 1989. ISBN 0-671-69098-1.
- Coborn, John. . New Jersey: TFH Publications. 1991. ISBN 978-0-86622-749-0.
- Cogger, Harold; Zweifel, Richard. . Sydney: Weldon Owen. 1992. ISBN 0-8317-2786-1.
- Conant, Roger; Collins, Joseph. . Boston, Massachusetts: Houghton Mifflin Company. 1991. ISBN 0-395-58389-6.
- Deane, John. . Whitefish, Montana: Kessinger Publishing. 1833: 412 [2012-09-01]. ISBN 1-56459-898-5. (原始内容存档于2019-10-09).
- Ditmars, Raymond L. . New York: E. R. Sanborn. 1906: 11.
- Ditmars, Raymond L. . New York: Macmillan. 1931: 11. ISBN 978-0-02-531730-7.
- Ditmars, Raymond L. . New York: Macmillan. 1933: 321.
- Ditmars, Raymond L; W. Bridges. . New York: D. Appleton and Company. 1935: 309.
- Ditmars, Raymond L. . Garden City, New York: Doubleday, Doran & Co. 1939: 305.
- Freiberg, Dr. Marcos; Walls, Jerry. . New Jersey: TFH Publications. 1984. ISBN 0-87666-567-9.
- Gibbons, J. Whitfield; Gibbons, Whit. . Alabama: University of Alabama Press. 1983: 164. ISBN 978-0-8173-0135-4.
- Mattison, Chris. . New Jersey: Princeton University Press. 2007: 272. ISBN 978-0-691-13295-2.
- McDiarmid, RW; Campbell, JA; Touré, T. 1. Herpetologists' League. 1999: 511. ISBN 1-893777-00-6.
- Mehrtens, John. . New York: Sterling. 1987. ISBN 0-8069-6461-8.
- Nóbrega Alves, RôMulo Romeu; Silva Vieira, Washington Luiz; Santana, Gindomar Gomes. . Biodiversity and Conservation. 2008, 17 (8): 2037–2049 [22 January 2009]. doi:10.1007/s10531-007-9305-0. (原始内容存档于2003-04-17).
- Romulus Whitaker (English edition); Tamil translation by O.Henry Francis. . National Book Trust. 1996. ISBN 81-237-1905-1.
- Rosenfeld, Arthur. . New York: Simon & Schuster. 1989: 293. ISBN 978-0-671-47654-0.
- Spawls, Steven; Branch, Bill. . Sanibel Island, Florida: Ralph Curtis Publishing. 1995: 192. ISBN 0-88359-029-8.
外部連結
- 台灣爬蟲資料庫:蛇
- (英文)整合分類學資訊系統:蛇類資料
- (英文)eNature:美國蛇類
- (英文)熱帶國家的蛇類
- (英文)印度大陸的蛇類
- (英文)歐洲蛇類 (页面存档备份,存于)
- (英文)毒蛇資料庫 (页面存档备份,存于)
- (英文)蛇類細胞:研究蛇類的組織 (页面存档备份,存于)
- (英文)蛇類世界
- (英文)「蛇是甚麼?」辨認蛇類的工具 (页面存档备份,存于)
- (英文)世界十大毒蛇排行榜
- (英文)瑞典自然歷史爬蟲學資料庫 (页面存档备份,存于)
- (日語)日本爬蟲兩棲類學會 (页面存档备份,存于)
- . Snake Track.com. [2012-09-01]. (原始内容存档于2019-05-10).
- . Biodiversity Heritage Library. [2012-09-01]. (原始内容存档于2021-05-06).
- . ITIS.
- . eNature. [2008-05-04]. (原始内容存档于2008-03-15).
- . Naturemagics Kerala Photo Gallery. [2008-05-04]. (原始内容存档于2009-03-21).
- . Snake Cell. [2012-09-01]. (原始内容存档于2020-12-19).(INDIA)
- . ReptilesWeb.com. [2008-05-04]. (原始内容存档于2008-07-03).
- . OPLIN. [2008-05-04]. (原始内容存档于2021-05-06).
- . Swedish Museum of Natural History. [2008-05-04]. (原始内容存档于2021-05-01).
- BBC Nature: (页面存档备份,存于) Snake news, and video clips from BBC programmes past and present.