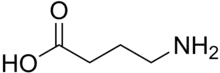
The GABA receptors are a class of receptors that respond to the neurotransmitter gamma-aminobutyric acid (GABA), the chief inhibitory compound in the mature vertebrate central nervous system. There are two classes of GABA receptors: GABAA and GABAB. GABAA receptors are ligand-gated ion channels (also known as ionotropic receptors); whereas GABAB receptors are G protein-coupled receptors, also called metabotropic receptors.
GABA receptors are neurotransmitters, a type of chemical messenger that exists in the nervous system. GABA has the function of reducing the excitation of neurons by blocking certain nerve transmissions. Thanks to the slowdown in message transmission, it helps to balance the mood and anxiety level. GABA calms the nervous system. Thus, excessive anxiety and stress are less common. If there are signal problems in GABA, it can cause mental health and nervous system disorders. For example, psychiatric and neurological problems may occur.[1]
GABA receptors are distributed in different areas of the brain, such as the cortex and basal ganglia. The cortex is the layer of the brain associated with seizure activity. For this reason, anti-seizure drugs work to increase the ratio of the reducing effect of the GABA receptor to the stimulating properties of another important neurotransmitter called glutamate. GABA is responsible for initiating the movements of muscle groups in the body and ensuring the continuation of movements. It is also responsible for sending signals to other areas of the brain. There are reasons for the increased activity of neurons because GABA activity is associated with anxiety, depression, focus and distraction problems.[2]
Ligand-gated ion channels
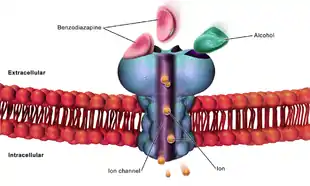
GABAA receptor
It has long been recognized that, for neurons that are stimulated by bicuculline and picrotoxin, the fast inhibitory response to GABA is due to direct activation of an anion channel.[3][4][5][6][7] This channel was subsequently termed the GABAA receptor.[8] Fast-responding GABA receptors are members of a family of Cys-loop ligand-gated ion channels.[9][10][11] Members of this superfamily, which includes nicotinic acetylcholine receptors, GABAA receptors, glycine and 5-HT3 receptors, possess a characteristic loop formed by a disulfide bond between two cysteine residues.[12]
In ionotropic GABAA receptors, binding of GABA molecules to their binding sites in the extracellular part of the receptor triggers opening of a chloride ion-selective pore.[13] The increased chloride conductance drives the membrane potential towards the reversal potential of the Cl¯ ion which is about –75 mV in neurons, inhibiting the firing of new action potentials. This mechanism is responsible for the sedative effects of GABAA allosteric agonists. In addition, activation of GABA receptors lead to the so-called shunting inhibition, which reduces the excitability of the cell independent of the changes in membrane potential.
There have been numerous reports of excitatory GABAA receptors. According to the excitatory GABA theory, this phenomenon is due to increased intracellular concentration of Cl¯ ions either during development of the nervous system[14][15] or in certain cell populations.[16][17][18] After this period of development, a chloride pump is upregulated and inserted into the cell membrane, pumping Cl− ions into the extracellular space of the tissue. Further openings via GABA binding to the receptor then produce inhibitory responses. Over-excitation of this receptor induces receptor remodeling and the eventual invagination of the GABA receptor. As a result, further GABA binding becomes inhibited and inhibitory postsynaptic potentials are no longer relevant.
However, the excitatory GABA theory has been questioned as potentially being an artefact of experimental conditions, with most data acquired in in-vitro brain slice experiments susceptible to un-physiological milieu such as deficient energy metabolism and neuronal damage. The controversy arose when a number of studies have shown that GABA in neonatal brain slices becomes inhibitory if glucose in perfusate is supplemented with ketone bodies, pyruvate, or lactate,[19][20] or that the excitatory GABA was an artefact of neuronal damage.[21] Subsequent studies from originators and proponents of the excitatory GABA theory have questioned these results,[22][23][24] but the truth remained elusive until the real effects of GABA could be reliably elucidated in intact living brain. Since then, using technology such as in-vivo electrophysiology/imaging and optogenetics, two in-vivo studies have reported the effect of GABA on neonatal brain, and both have shown that GABA is indeed overall inhibitory, with its activation in the developing rodent brain not resulting in network activation,[25] and instead leading to a decrease of activity.[26][27]
GABA receptors influence neural function by coordinating with glutamatergic processes.[28] Inhibitory postsynaptic potential refers to the local hyperpolarization created in a neuron by the activation of the synapse made by a nerve ending. GABAA is also the fast component. They are ionotropic receptors. It provides selective permeability to Cl- channels. In addition, GABAA subunits are also very clinically important. [29]
GABAA-ρ receptor
A subclass of ionotropic GABA receptors, insensitive to typical allosteric modulators of GABAA receptor channels such as benzodiazepines and barbiturates,[30][31][32] was designated GABAС receptor.[33][34] Native responses of the GABAC receptor type occur in retinal bipolar or horizontal cells across vertebrate species.[35][36][37][38]
GABAС receptors are exclusively composed of ρ (rho) subunits that are related to GABAA receptor subunits.[39][40][41] Although the term "GABAС receptor" is frequently used, GABAС may be viewed as a variant within the GABAA receptor family.[9] Others have argued that the differences between GABAС and GABAA receptors are large enough to justify maintaining the distinction between these two subclasses of GABA receptors.[42][43] However, since GABAС receptors are closely related in sequence, structure, and function to GABAA receptors and since other GABAA receptors besides those containing ρ subunits appear to exhibit GABAС pharmacology, the Nomenclature Committee of the IUPHAR has recommended that the GABAС term no longer be used and these ρ receptors should be designated as the ρ subfamily of the GABAA receptors (GABAA-ρ).[44]
G protein-coupled receptors
GABAB receptor
A slow response to GABA is mediated by GABAB receptors,[45] originally defined on the basis of pharmacological properties.[46]
In studies focused on the control of neurotransmitter release, it was noted that a GABA receptor was responsible for modulating evoked release in a variety of isolated tissue preparations. This ability of GABA to inhibit neurotransmitter release from these preparations was not blocked by bicuculline, was not mimicked by isoguvacine, and was not dependent on Cl¯, all of which are characteristic of the GABAA receptor. The most striking discovery was the finding that baclofen (β-parachlorophenyl GABA), a clinically employed muscle relaxant[47][48] mimicked, in a stereoselective manner, the effect of GABA.
Later ligand-binding studies provided direct evidence of binding sites for baclofen on central neuronal membranes.[49] cDNA cloning confirmed that the GABAB receptor belongs to the family of G-protein coupled receptors.[50] Additional information on GABAB receptors has been reviewed elsewhere.[51][52][53][54][55][56][57][58]
GABAB Inhibitor is the slow component of the postsynaptic potential. They are metabotropic receptors. It provides Ca+2 channel inhibition or K+ channel activation. The inhibitory postsynaptic potential is long lasting and occurs slowly. It also inhibits neurotransmitter release by inhibiting Ca+2 Channels. It also inhibits adenylate cyclase and cAMP formation due to its docking with ion channels.[59]
GABA receptor gene polymorphisms
Two separate genes on two chromosomes control GABA synthesis - glutamate decarboxylase and alpha-ketoglutarate decarboxylase genes - though not much research has been done to explain this polygenic phenomenon.[60] GABA receptor genes have been studied more in depth, and many have hypothesized about the deleterious effects of polymorphisms in these receptor genes. The most common single nucleotide polymorphisms (SNPs) occurring in GABA receptor genes rho 1, 2, and 3 (GABBR1, GABBR2, and GABBR3) have been more recently explored in literature, in addition to the potential effects of these polymorphisms. However, some research has demonstrated that there is evidence that these polymorphisms caused by single base pair variations may be harmful.
It was discovered that the minor allele of a single nucleotide polymorphism at GABBR1 known as rs1186902 is significantly associated with a later age of onset for migraines,[61] but for the other SNPs, no differences were discovered between genetic and allelic variations in the control vs. migraine participants. Similarly, in a study examining SNPs in rho 1, 2, and 3, and their implication in essential tremor, a nervous system disorder, it was discovered that there were no differences in the frequencies of the allelic variants of polymorphisms for control vs. essential tremor participants.[62] On the other hand, research examining the effect of SNPs in participants with restless leg syndrome found an "association between GABRR3rs832032 polymorphism and the risk for RLS, and a modifier effect of GABRA4 rs2229940 on the age of onset of RLS" - the latter of which is a modifier gene polymorphism.[63] The most common GABA receptor SNPs do not correlate with deleterious health effects in many cases, but do in a few.
One significant example of a deleterious mutation is the major association between several GABA receptor gene polymorphisms and schizophrenia. Because GABA is integral to the release of inhibitory neurotransmitters which produce a calming effect and play a role in reducing anxiety, stress, and fear, it is not surprising that polymorphisms in these genes result in more consequences relating to mental health than to physical health. Of an analysis on 19 SNPs on various GABA receptor genes, five SNPs in the GABBR2 group were found to be significantly associated with schizophrenia,[64] which produce the unexpected haplotype frequencies not found in the studies mentioned previously.
Several studies have verified association between alcohol use disorder and the rs279858 polymorphism on the GABRA2 gene e, and higher negative alcohol effects scores for individuals who were homozygous at six SNPs.[65] Furthermore, a study examining polymorphisms in the GABA receptor beta 2 subunit gene found an association with schizophrenia and bipolar disorder, and examined three SNPs and their effects on disease frequency and treatment dosage.[66] A major finding of this study was that functional psychosis should be conceptualized as a scale of phenotypes rather than distinct categories.
See also
References
- ↑ Menzikov SA, Zaichenko DM, Moskovtsev AA, Morozov SG, Kubatiev AA. Physiological Role of ATPase for GABAA Receptor Resensitization. Int J Mol Sci. 2022 May 10;23(10):5320. doi: 10.3390/ijms23105320. PMID: 35628132; PMCID: PMC9141714.
- ↑ Sinha R, Grimes WN, Wallin J, Ebbinghaus BN, Luu K, Cherry T, Rieke F, Rudolph U, Wong RO, Hoon M. Transient expression of a GABA receptor subunit during early development is critical for inhibitory synapse maturation and function. Curr Biol. 2021 Oct 11;31(19):4314-4326.e5. doi: 10.1016/j.cub.2021.07.059. Epub 2021 Aug 24. PMID: 34433078; PMCID: PMC8511107.
- ↑ Kuffler SW, Edwards C (November 1958). "Mechanism of gamma aminobutyric acid (GABA) action and its relation to synaptic inhibition". Journal of Neurophysiology. 21 (6): 589–610. doi:10.1152/jn.1958.21.6.589. PMID 13599049. Archived from the original on 2004-08-03.
- ↑ Kravitz EA, Kuffler SW, Potter DD (September 1963). "Gamma-Aminobutyric Acid and Other Blocking Compounds in Crustacea: III. Their Relative Concentrations in Separated Motor and Inhibitory Axons". Journal of Neurophysiology. 26 (5): 739–51. doi:10.1152/jn.1963.26.5.739. PMID 14065325.
- ↑ Krnjević K, Schwartz S (1967). "The action of gamma-aminobutyric acid on cortical neurones". Experimental Brain Research. 3 (4): 320–36. doi:10.1007/BF00237558. PMID 6031164. S2CID 6891616.
- ↑ Takeuchi A, Takeuchi N (August 1967). "Anion permeability of the inhibitory post-synaptic membrane of the crayfish neuromuscular junction". The Journal of Physiology. 191 (3): 575–90. doi:10.1113/jphysiol.1967.sp008269. PMC 1365493. PMID 6051794.
- ↑ Takeuchi A, Takeuchi N (November 1969). "A study of the action of picrotoxin on the inhibitory neuromuscular junction of the crayfish". The Journal of Physiology. 205 (2): 377–91. doi:10.1113/jphysiol.1969.sp008972. PMC 1348609. PMID 5357245.
- ↑ Takeuchi A, Onodera K (March 1972). "Effect of bicuculline on the GABA receptor of the crayfish neuromuscular junction". Nature. 236 (63): 55–6. doi:10.1038/236055a0. PMID 4502428. S2CID 12978932.
- 1 2 Barnard EA, Skolnick P, Olsen RW, Mohler H, Sieghart W, Biggio G, et al. (June 1998). "International Union of Pharmacology. XV. Subtypes of gamma-aminobutyric acidA receptors: classification on the basis of subunit structure and receptor function". Pharmacological Reviews. 50 (2): 291–313. PMID 9647870.
- ↑ Hevers W, Lüddens H (August 1998). "The diversity of GABAA receptors. Pharmacological and electrophysiological properties of GABAA channel subtypes". Molecular Neurobiology. 18 (1): 35–86. doi:10.1007/BF02741459. PMID 9824848. S2CID 32359279.
- ↑ Sieghart W, Sperk G (August 2002). "Subunit composition, distribution and function of GABA(A) receptor subtypes". Current Topics in Medicinal Chemistry. 2 (8): 795–816. doi:10.2174/1568026023393507. PMID 12171572.
- ↑ Phulera S, Zhu H, Yu J, Claxton DP, Yoder N, Yoshioka C, Gouaux E (July 2018). "A receptor in complex with GABA". eLife. 7: e39383. doi:10.7554/eLife.39383. PMC 6086659. PMID 30044221.
- ↑ Phulera S, Zhu H, Yu J, Claxton DP, Yoder N, Yoshioka C, Gouaux E (July 2018). "A receptor in complex with GABA". eLife. 7: e39383. doi:10.7554/eLife.39383. PMC 6086659. PMID 30044221.
- ↑ Ben-Ari Y, Khazipov R, Leinekugel X, Caillard O, Gaiarsa JL (November 1997). "GABAA, NMDA and AMPA receptors: a developmentally regulated 'ménage à trois'". Trends Neurosci. 20 (11): 523–9. doi:10.1016/S0166-2236(97)01147-8. PMID 9364667. S2CID 8022055.
- ↑ Taketo M, Yoshioka T (2000). "Developmental change of GABA(A) receptor-mediated current in rat hippocampus". Neuroscience. 96 (3): 507–14. doi:10.1016/S0306-4522(99)00574-6. PMID 10717431. S2CID 22103661.
- ↑ Tomiko SA, Taraskevich PS, Douglas WW (February 1983). "GABA acts directly on cells of pituitary pars intermedia to alter hormone output". Nature. 301 (5902): 706–7. Bibcode:1983Natur.301..706T. doi:10.1038/301706a0. PMID 6828152. S2CID 4326183.
- ↑ Cherubini E, Gaiarsa JL, Ben-Ari Y (December 1991). "GABA: an excitatory transmitter in early postnatal life". Trends Neurosci. 14 (12): 515–9. doi:10.1016/0166-2236(91)90003-D. PMID 1726341. S2CID 3971981.
- ↑ Lamsa K, Taira T (September 2003). "Use-dependent shift from inhibitory to excitatory GABAA receptor action in SP-O interneurons in the rat hippocampal CA3 area". J. Neurophysiol. 90 (3): 1983–95. doi:10.1152/jn.00060.2003. PMID 12750426. S2CID 17650510.
- ↑ Rheims S, Holmgren CD, Chazal G, Mulder J, Harkany T, Zilberter T, Zilberter Y (August 2009). "GABA action in immature neocortical neurons directly depends on the availability of ketone bodies". Journal of Neurochemistry. 110 (4): 1330–8. doi:10.1111/j.1471-4159.2009.06230.x. PMID 19558450.
- ↑ Holmgren CD, Mukhtarov M, Malkov AE, Popova IY, Bregestovski P, Zilberter Y (February 2010). "Energy substrate availability as a determinant of neuronal resting potential, GABA signaling and spontaneous network activity in the neonatal cortex in vitro". Journal of Neurochemistry. 112 (4): 900–12. doi:10.1111/j.1471-4159.2009.06506.x. PMID 19943846.
- ↑ Dzhala V, Valeeva G, Glykys J, Khazipov R, Staley K (March 2012). "Traumatic alterations in GABA signaling disrupt hippocampal network activity in the developing brain". The Journal of Neuroscience. 32 (12): 4017–31. doi:10.1523/JNEUROSCI.5139-11.2012. PMC 3333790. PMID 22442068.
- ↑ Kirmse K, Witte OW, Holthoff K (November 2010). "GABA depolarizes immature neocortical neurons in the presence of the ketone body β-hydroxybutyrate". The Journal of Neuroscience. 30 (47): 16002–7. doi:10.1523/JNEUROSCI.2534-10.2010. PMC 6633760. PMID 21106838.
- ↑ Ruusuvuori E, Kirilkin I, Pandya N, Kaila K (November 2010). "Spontaneous network events driven by depolarizing GABA action in neonatal hippocampal slices are not attributable to deficient mitochondrial energy metabolism". The Journal of Neuroscience. 30 (46): 15638–42. doi:10.1523/JNEUROSCI.3355-10.2010. PMC 6633692. PMID 21084619.
- ↑ Tyzio R, Allene C, Nardou R, Picardo MA, Yamamoto S, Sivakumaran S, et al. (January 2011). "Depolarizing actions of GABA in immature neurons depend neither on ketone bodies nor on pyruvate". The Journal of Neuroscience. 31 (1): 34–45. doi:10.1523/JNEUROSCI.3314-10.2011. PMC 6622726. PMID 21209187.
- ↑ Kirmse K, Kummer M, Kovalchuk Y, Witte OW, Garaschuk O, Holthoff K (July 2015). "GABA depolarizes immature neurons and inhibits network activity in the neonatal neocortex in vivo". Nature Communications. 6: 7750. Bibcode:2015NatCo...6.7750K. doi:10.1038/ncomms8750. PMID 26177896.
- ↑ Valeeva G, Tressard T, Mukhtarov M, Baude A, Khazipov R (June 2016). "An Optogenetic Approach for Investigation of Excitatory and Inhibitory Network GABA Actions in Mice Expressing Channelrhodopsin-2 in GABAergic Neurons". The Journal of Neuroscience. 36 (22): 5961–73. doi:10.1523/JNEUROSCI.3482-15.2016. PMC 6601813. PMID 27251618.
- ↑ Zilberter M (October 2016). "Reality of Inhibitory GABA in Neonatal Brain: Time to Rewrite the Textbooks?". The Journal of Neuroscience. 36 (40): 10242–10244. doi:10.1523/JNEUROSCI.2270-16.2016. PMC 6705588. PMID 27707962.
- ↑ Farahmandfar M, Akbarabadi A, Bakhtazad A, Zarrindast MR (March 2017). "Recovery from ketamine-induced amnesia by blockade of GABA-A receptor in the medial prefrontal cortex of mice". Neuroscience. 344: 48–55. doi:10.1016/j.neuroscience.2016.02.056. PMID 26944606. S2CID 24077379.
- ↑ Marques TR, Ashok AH, Angelescu I, Borgan F, Myers J, Lingford-Hughes A, Nutt DJ, Veronese M, Turkheimer FE, Howes OD. GABA-A receptor differences in schizophrenia: a positron emission tomography study using [11C]Ro154513. Mol Psychiatry. 2021 Jun;26(6):2616-2625. doi: 10.1038/s41380-020-0711-y. Epub 2020 Apr 15. PMID: 32296127; PMCID: PMC8440185.
- ↑ Sivilotti L, Nistri A (1991). "GABA receptor mechanisms in the central nervous system". Prog. Neurobiol. 36 (1): 35–92. doi:10.1016/0301-0082(91)90036-Z. PMID 1847747. S2CID 31732465.
- ↑ Bormann J, Feigenspan A (December 1995). "GABAC receptors". Trends Neurosci. 18 (12): 515–9. doi:10.1016/0166-2236(95)98370-E. PMID 8638289. S2CID 40853254.
- ↑ Johnston GA (September 1996). "GABAc receptors: relatively simple transmitter -gated ion channels?". Trends Pharmacol. Sci. 17 (9): 319–23. doi:10.1016/0165-6147(96)10038-9. PMID 8885697.
- ↑ Drew CA, Johnston GA, Weatherby RP (December 1984). "Bicuculline-insensitive GABA receptors: studies on the binding of (-)-baclofen to rat cerebellar membranes". Neurosci. Lett. 52 (3): 317–21. doi:10.1016/0304-3940(84)90181-2. PMID 6097844. S2CID 966075.
- ↑ Zhang D, Pan ZH, Awobuluyi M, Lipton SA (March 2001). "Structure and function of GABA(C) receptors: a comparison of native versus recombinant receptors". Trends Pharmacol. Sci. 22 (3): 121–32. doi:10.1016/S0165-6147(00)01625-4. PMID 11239575.
- ↑ Feigenspan A, Wässle H, Bormann J (January 1993). "Pharmacology of GABA receptor Cl- channels in rat retinal bipolar cells". Nature. 361 (6408): 159–62. Bibcode:1993Natur.361..159F. doi:10.1038/361159a0. PMID 7678450. S2CID 4347233.
- ↑ Qian H, Dowling JE (January 1993). "Novel GABA responses from rod-driven retinal horizontal cells". Nature. 361 (6408): 162–4. Bibcode:1993Natur.361..162Q. doi:10.1038/361162a0. PMID 8421521. S2CID 4320616.
- ↑ Lukasiewicz PD (June 1996). "GABAC receptors in the vertebrate retina". Mol. Neurobiol. 12 (3): 181–94. doi:10.1007/BF02755587. PMID 8884747. S2CID 37167159.
- ↑ Wegelius K, Pasternack M, Hiltunen JO, Rivera C, Kaila K, Saarma M, Reeben M (January 1998). "Distribution of GABA receptor rho subunit transcripts in the rat brain". Eur. J. Neurosci. 10 (1): 350–7. doi:10.1046/j.1460-9568.1998.00023.x. PMID 9753143. S2CID 25863134.
- ↑ Shimada S, Cutting G, Uhl GR (April 1992). "gamma-Aminobutyric acid A or C receptor? gamma-Aminobutyric acid rho 1 receptor RNA induces bicuculline-, barbiturate-, and benzodiazepine-insensitive gamma-aminobutyric acid responses in Xenopus oocytes". Mol. Pharmacol. 41 (4): 683–7. PMID 1314944.
- ↑ Kusama T, Spivak CE, Whiting P, Dawson VL, Schaeffer JC, Uhl GR (May 1993). "Pharmacology of GABA rho 1 and GABA alpha/beta receptors expressed in Xenopus oocytes and COS cells". Br. J. Pharmacol. 109 (1): 200–6. doi:10.1111/j.1476-5381.1993.tb13554.x. PMC 2175610. PMID 8388298.
- ↑ Kusama T, Wang TL, Guggino WB, Cutting GR, Uhl GR (March 1993). "GABA rho 2 receptor pharmacological profile: GABA recognition site similarities to rho 1". Eur. J. Pharmacol. 245 (1): 83–4. doi:10.1016/0922-4106(93)90174-8. PMID 8386671.
- ↑ Chebib M, Johnston GA (April 2000). "GABA-Activated ligand gated ion channels: medicinal chemistry and molecular biology". J. Med. Chem. 43 (8): 1427–47. doi:10.1021/jm9904349. PMID 10780899.
- ↑ Bormann J (January 2000). "The 'ABC' of GABA receptors". Trends Pharmacol. Sci. 21 (1): 16–9. doi:10.1016/S0165-6147(99)01413-3. PMID 10637650.
- ↑ Olsen RW, Sieghart W (September 2008). "International Union of Pharmacology. LXX. Subtypes of γ-Aminobutyric AcidA Receptors: Classification on the Basis of Subunit Composition, Pharmacology, and Function. Update". Pharmacological Reviews. 60 (3): 243–60. doi:10.1124/pr.108.00505. PMC 2847512. PMID 18790874.
- ↑ Bowery NG, Bettler B, Froestl W, Gallagher JP, Marshall F, Raiteri M, Bonner TI, Enna SJ (June 2002). "International Union of Pharmacology. XXXIII. Mammalian gamma-aminobutyric acid(B) receptors: structure and function". Pharmacological Reviews. 54 (2): 247–64. doi:10.1124/pr.54.2.247. PMID 12037141. S2CID 86015084.
- ↑ Bowery NG, Hill DR, Hudson AL, Doble A, Middlemiss DN, Shaw J, Turnbull M (January 1980). "(-)Baclofen decreases neurotransmitter release in the mammalian CNS by an action at a novel GABA receptor". Nature. 283 (5742): 92–4. Bibcode:1980Natur.283...92B. doi:10.1038/283092a0. PMID 6243177. S2CID 4238700.
- ↑ Bein HJ (1972). "Pharmacological differentiations of muscle relaxants". In Birkmayer W (ed.). Spasticity: A Topical Survey. Hans Hubert Bern, Switzerland. pp. 76–89. ISBN 3-456-00390-0.
- ↑ Keberle H, Faigle JW (1972). "Synthesis and structure-activity relationship of the gamma-aminobutyric acid derivatives". In Birkmayer W (ed.). Spasticity: A Topical Survey. Hans Hubert Bern, Switzerland. pp. 76–89. ISBN 3-456-00390-0.
- ↑ Hill DR, Bowery NG (March 1981). "3H-baclofen and 3H-GABA bind to bicuculline-insensitive GABA B sites in rat brain". Nature. 290 (5802): 149–52. Bibcode:1981Natur.290..149H. doi:10.1038/290149a0. PMID 6259535. S2CID 4335907.
- ↑ Kaupmann K, Huggel K, Heid J, Flor PJ, Bischoff S, Mickel SJ, McMaster G, Angst C, Bittiger H, Froestl W, Bettler B (March 1997). "Expression cloning of GABA(B) receptors uncovers similarity to metabotropic glutamate receptors". Nature. 386 (6622): 239–46. Bibcode:1997Natur.386..239K. doi:10.1038/386239a0. PMID 9069281. S2CID 4345443.
- ↑ Enna SJ (October 1997). "GABAB receptor agonists and antagonists: pharmacological properties and therapeutic possibilities". Expert Opin Investig Drugs. 6 (10): 1319–25. doi:10.1517/13543784.6.10.1319. PMID 15989503.
- ↑ Bowery, N. G.; Enna, S. J. (1997). The GABA receptors. Totowa, NJ: Humana Press. ISBN 0-89603-458-5.
- ↑ Kaupmann K, Malitschek B, Schuler V, Heid J, Froestl W, Beck P, Mosbacher J, Bischoff S, Kulik A, Shigemoto R, Karschin A, Bettler B (December 1998). "GABA(B)-receptor subtypes assemble into functional heteromeric complexes". Nature. 396 (6712): 683–7. Bibcode:1998Natur.396..683K. doi:10.1038/25360. PMID 9872317. S2CID 4421681.
- ↑ Kaupmann K, Schuler V, Mosbacher J, Bischoff S, Bittiger H, Heid J, Froestl W, Leonhard S, Pfaff T, Karschin A, Bettler B (December 1998). "Human γ-aminobutyric acid type B receptors are differentially expressed and regulate inwardly rectifying K+ channels". Proc. Natl. Acad. Sci. U.S.A. 95 (25): 14991–6. Bibcode:1998PNAS...9514991K. doi:10.1073/pnas.95.25.14991. PMC 24563. PMID 9844003.
- ↑ Marshall FH, Jones KA, Kaupmann K, Bettler B (October 1999). "GABA receptors - the first 7TM heterodimers". Trends Pharmacol. Sci. 20 (10): 396–9. doi:10.1016/S0165-6147(99)01383-8. PMID 10498952.
- ↑ Marshall FH, White J, Main M, Green A, Wise A (August 1999). "GABA(B) receptors function as heterodimers". Biochem. Soc. Trans. 27 (4): 530–5. doi:10.1042/bst0270530. PMID 10917635.
- ↑ Bowery NG, Enna SJ (January 2000). "gamma-aminobutyric acid(B) receptors: first of the functional metabotropic heterodimers". J. Pharmacol. Exp. Ther. 292 (1): 2–7. PMID 10604925.
- ↑ Enna SJ (2001). "GABAB receptor signaling pathways". In Möhler H (ed.). Pharmacology of GABA and Glycine Neurotransmission. Handbook of Experimental Pharmacology. Vol. 150. Berlin: Springer. pp. 329–342. ISBN 3-540-67616-3.
- ↑ Tomita S. Molecular constituents and localization of the ionotropic GABA receptor complex in vivo. Curr Opin Neurobiol. 2019 Aug;57:81-86. doi: 10.1016/j.conb.2019.01.017. Epub 2019 Feb 19. PMID: 30784980; PMCID: PMC6629498.
- ↑ Kanwal, Simab; Incharoensakdi, Aran (2020-01-01). "GABA synthesis mediated by γ-aminobutanal dehydrogenase in Synechocystis sp. PCC6803 with disrupted glutamate and α-ketoglutarate decarboxylase genes". Plant Science. 290: 110287. doi:10.1016/j.plantsci.2019.110287. ISSN 0168-9452. PMID 31779897. S2CID 204162907.
- ↑ García-Martín, Elena; Martínez, Carmen; Serrador, Mercedes; Alonso-Navarro, Hortensia; Navacerrada, Francisco; Esguevillas, Gara; García-Albea, Esteban; Agúndez, José A. G.; Jiménez-Jiménez, Félix Javier (2017). "Gamma‐Aminobutyric Acid (Gaba) Receptors Rho (Gabrr) Gene Polymorphisms and Risk for Migraine". Headache: The Journal of Head and Face Pain. 57 (7): 1118–1135. doi:10.1111/head.13122. PMID 28699326. S2CID 12303665.
- ↑ García-Martín, Elena; Martínez, Carmen; Alonso-Navarro, Hortensia; Benito-León, Julián; Lorenzo-Betancor, Oswaldo; Pastor, Pau; Puertas, Inmaculada; Rubio, Lluisa; López-Alburquerque, Tomás; Agúndez, José A. G.; Jiménez-Jiménez, Félix Javier (2011). "Gamma-aminobutyric acid (GABA) receptor rho (GABRR) polymorphisms and risk for essential tremor". Journal of Neurology. 258 (2): 203–211. doi:10.1007/s00415-010-5708-z. PMID 20820800. S2CID 22082250.
- ↑ Jiménez-Jiménez, Félix Javier; Esguevillas, Gara; Alonso-Navarro, Hortensia; Zurdo, Martín; Turpín-Fenoll, Laura; Millán-Pascual, Jorge; Adeva-Bartolomé, Teresa; Cubo, Esther; Navacerrada, Francisco; Amo, Gemma; Rojo-Sebastián, Ana; Rubio, Lluisa; Díez-Fairén, Mónica; Pastor, Pau; Calleja, Marisol; Plaza-Nieto, José Francisco; Pilo-De-La-Fuente, Belén; Arroyo-Solera, Margarita; García-Albea, Esteban; Agúndez, José A. G.; García-Martín, Elena (2018). "Gamma-aminobutyric acid (GABA) receptors genes polymorphisms and risk for restless legs syndrome". The Pharmacogenomics Journal. 18 (4): 565–577. doi:10.1038/s41397-018-0023-7. PMID 29720720. S2CID 13756330.
- ↑ Lo, W.-S.; Lau, C.-F.; Xuan, Z.; Chan, C.-F.; Feng, G.-Y.; He, L.; Cao, Z.-C.; Liu, H.; Luan, Q.-M.; Xue, H. (June 2004). "Association of SNPs and haplotypes in GABA A receptor β 2 gene with schizophrenia". Molecular Psychiatry. 9 (6): 603–608. doi:10.1038/sj.mp.4001461. ISSN 1476-5578. PMID 14699426. S2CID 5567422.
- ↑ Koulentaki, Mairi; Kouroumalis, Elias (2018-06-01). "GABAA receptor polymorphisms in alcohol use disorder in the GWAS era". Psychopharmacology. 235 (6): 1845–1865. doi:10.1007/s00213-018-4918-4. ISSN 1432-2072. PMID 29721579. S2CID 13744792.
- ↑ Chen, Jianhuan; Tsang, Shui-Ying; Zhao, Cun-You; Pun, Frank W.; Yu, Zhiliang; Mei, Lingling; Lo, Wing-Sze; Fang, Shisong; Liu, Hua; Stöber, Gerald; Xue, Hong (2009-12-01). "GABRB2 in schizophrenia and bipolar disorder: disease association, gene expression and clinical correlations". Biochemical Society Transactions. 37 (6): 1415–1418. doi:10.1042/BST0371415. ISSN 0300-5127. PMID 19909288. S2CID 10742771.
External links
- IUPHAR GPCR Database - GABAB receptors
- GABA+Receptor at the U.S. National Library of Medicine Medical Subject Headings (MeSH)