Light-gated ion channels are a family of ion channels regulated by electromagnetic radiation. Other gating mechanisms for ion channels include voltage-gated ion channels, ligand-gated ion channels, mechanosensitive ion channels, and temperature-gated ion channels. Most light-gated ion channels have been synthesized in the laboratory for study, although two naturally occurring examples, channelrhodopsin and anion-conducting channelrhodopsin, are currently known.[1][2] Photoreceptor proteins, which act in a similar manner to light-gated ion channels, are generally classified instead as G protein-coupled receptors.
Mechanism
Light-gated ion channels function in a similar manner to other gated ion channels. Such transmembrane proteins form pores through lipid bilayers to facilitate the passage of ions. These ions move from one side of the membrane to another under the influence of an electrochemical gradient. When exposed to a stimulus, a conformational change occurs in the transmembrane region of the protein to open or close the ion channel. In the specific case of light-gated ion channels, the transmembrane proteins are usually coupled with a smaller molecule that acts as a photoswitch, whereby photons bind to the switching molecule, to then alter the conformation of the proteins, so that the pore changes from a closed state to an open state, or vice versa, thereby increasing or decreasing ion conductance. Retinal is a good example of a molecular photoswitch and is found in the naturally occurring channelrhodopsins.[3][4]
Synthetic isoforms
Once channelrhosopsin had been identified and characterized, the channel's ion selectivity was modified in order to control membrane potential through optogenetic control. Directed mutations of the channel changed the charges lining the pore, resulting in a pore which instead excluded cations in favor of anions.[5]
Other types of gated ion channels, ligand-gated and voltage-gated, have been synthesized with a light-gated component in an attempt to better understand their nature and properties. By the addition of a light-gated section, the kinetics and mechanisms of operation can be studied in depth. For example, the addition of a light-gated component allows for the introduction of many highly similar ligands to be introduced to the binding site of a ligand-gated ion channel to assist in the determination of the mechanism.
Such ion channels have been modified by binding a photoswitch to confer photosensitivity on the ion channel. This is done through careful selection of a tether which can lengthen or shorten through photoisomerization. One side of the tether is bound to the ion channel protein and the other end of the tether is bound to a blocking group, which has a high binding affinity for an exposed portion of the pore. When the tether is lengthened, it allows the blocking section to bind to the pore and prevent ionic current. When the tether is shortened, it disrupts this obstruction and opens the pore. Kinetic studies have demonstrated that fine temporal and spatial control can be achieved in this manner.[6][7]
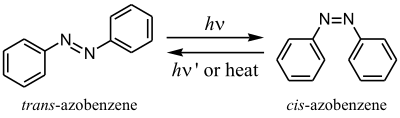
Azobenzene is a common choice for the functional portion of a tether for synthetically-developed light-gated ion channels because of its well documented length change as either cis or trans isomers, as well as the excitation wavelength needed to induce photoisomerization. Azobenzene converts to its longer trans-isomer at a wavelength of λ=500 nm and to its cis-isomer at λ=380 nm.[6]
In 1980, the first ion channel to be adapted for study with a light-gated mechanism was the nicotinic acetylcholine receptor.[8] This receptor was well-known at the time, and so was aptly suited to adaptation, and allowed for a study of the kinetics as not allowed before.
The expression of light-gated ion channels in a specific cell type through promoter control allows for the regulation of cell potential by either depolarizing the membrane to 0 mV for cation-permeant channelrhodopsin or by holding the voltage at –67 mV for anion-conducting channelrhodopsin.[9] Depolarization can conduct a current in the range of 5 fA per channel and occurs on the timescale of action potentials and neurotransmitter exocytosis.[10][4] They have an advantage over other types of ion channel regulation in that they provide non-invasive, reversible membrane potential changes with fine temporal and spatial control granted by induction through laser stimuli.[3][6] They reliably stimulate single action potentials with rapid depolarization and can be utilized in vivo because they do not require high intensity illumination to maintain function, unlike other techniques like light-activated proton pumps and photoactivatable probes.[5][10]
Examples
Examples of light-gated ion channels occur in both natural and synthetic environments. These include:
Naturally occurring
- Channelrhodopsins were the first discovered family of light-gated ion channels.
Synthetically adapted
- Nicotinic acetylcholine receptor was the first ion channel to be synthetically adapted with a light-gated mechanism.
- Light-activated potassium channels have been engineered from bacterial K+ channels with the goal of inhibiting neuronal activity upon illumination.[11] A second strategy is to combine a cyclic nucleotide gated K+ channel with a photoactivated adenylyl cyclase to inhibit neuronal activity at very low light levels.[12][13]
- Many other fully synthetic, light-gated channels have been produced as well.[14][15]
References
- ↑ "Engineering Light-Gated Ion Channels" — Biochemistry, 45 (51), 15129 -15141, 2006
- ↑ Govorunova, Elena G.; Sineshchekov, Oleg A.; Janz, Roger; Liu, Xiaoqin; Spudich, John L. (2015-08-07). "Natural light-gated anion channels: A family of microbial rhodopsins for advanced optogenetics". Science. 349 (6248): 647–650. doi:10.1126/science.aaa7484. ISSN 0036-8075. PMC 4764398. PMID 26113638.
- 1 2 Nagel, Georg; Brauner, Martin; Liewald, Jana F.; Adeishvili, Nona; Bamberg, Ernst; Gottschalk, Alexander (2005). "Light Activation of Channelrhodopsin-2 in Excitable Cells of Caenorhabditis elegans Triggers Rapid Behavioral Responses". Current Biology. 15 (24): 2279–2284. doi:10.1016/j.cub.2005.11.032. PMID 16360690.
- 1 2 Nagel, Georg; Szellas, Tanjef; Huhn, Wolfram; Kateriya, Suneel; Adeishvili, Nona; Berthold, Peter; Ollig, Doris; Hegemann, Peter; Bamberg, Ernst (2003-11-25). "Channelrhodopsin-2, a directly light-gated cation-selective membrane channel". Proceedings of the National Academy of Sciences. 100 (24): 13940–13945. doi:10.1073/pnas.1936192100. ISSN 0027-8424. PMC 283525. PMID 14615590.
- 1 2 Wietek, Jonas; Wiegert, J. Simon; Adeishvili, Nona; Schneider, Franziska; Watanabe, Hiroshi; Tsunoda, Satoshi P.; Vogt, Arend; Elstner, Marcus; Oertner, Thomas G. (2014-04-25). "Conversion of Channelrhodopsin into a Light-Gated Chloride Channel". Science. 344 (6182): 409–412. doi:10.1126/science.1249375. ISSN 0036-8075. PMID 24674867. S2CID 206554245.
- 1 2 3 Banghart, Matthew; Borges, Katharine; Isacoff, Ehud; Trauner, Dirk; Kramer, Richard H (December 2004). "Light-activated ion channels for remote control of neuronal firing". Nature Neuroscience. 7 (12): 1381–1386. doi:10.1038/nn1356. ISSN 1546-1726. PMC 1447674. PMID 15558062.
- ↑ Jog, Parag V.; Gin, Mary S. (2008-09-01). "A Light-Gated Synthetic Ion Channel". Organic Letters. 10 (17): 3693–3696. doi:10.1021/ol8013045. ISSN 1523-7060. PMID 18656946.
- ↑ "A covalently bound photoisomerizable agonist. Comparison with reversibly bound agonists at electrophorus electroplaques" — The Journal of General Physiology, Vol 75, 207-232
- ↑ Berndt, Andre; Lee, Soo Yeun; Ramakrishnan, Charu; Deisseroth, Karl (2014-04-25). "Structure-Guided Transformation of Channelrhodopsin into a Light-Activated Chloride Channel". Science. 344 (6182): 420–424. doi:10.1126/science.1252367. ISSN 0036-8075. PMC 4096039. PMID 24763591.
- 1 2 Ishizuka, Toru; Kakuda, Masaaki; Araki, Rikita; Yawo, Hiromu (2006). "Kinetic evaluation of photosensitivity in genetically engineered neurons expressing green algae light-gated channels". Neuroscience Research. 54 (2): 85–94. doi:10.1016/j.neures.2005.10.009. PMID 16298005. S2CID 17576414.
- ↑ Cosentino, C.; Alberio, L.; Gazzarrini, S.; Aquila, M.; Romano, E.; Cermenati, S.; Zuccolini, P.; Petersen, J.; Beltrame, M.; Etten, J. L. Van; Christie, J. M.; Thiel, G.; Moroni, A. (2015). "Engineering of a light-gated potassium channel". Science. 348 (6235): 707–710. doi:10.1126/science.aaa2787. PMID 25954011. S2CID 27853990.
- ↑ Beck, Sebastian; Yu-Strzelczyk, Jing; Pauls, Dennis; Constantin, Oana M.; Gee, Christine E.; Ehmann, Nadine; Kittel, Robert J.; Nagel, Georg; Gao, Shiqiang (2018). "Synthetic Light-Activated Ion Channels for Optogenetic Activation and Inhibition". Frontiers in Neuroscience. 12: 643. doi:10.3389/fnins.2018.00643. ISSN 1662-453X. PMC 6176052. PMID 30333716.
- ↑ Bernal Sierra, Yinth Andrea; Rost, Benjamin R.; Pofahl, Martin; Fernandes, António Miguel; Kopton, Ramona A.; Moser, Sylvain; Holtkamp, Dominik; Masala, Nicola; Beed, Prateep; Tukker, John J.; Oldani, Silvia (2018). "Potassium channel-based optogenetic silencing". Nature Communications. 9 (1): 4611. doi:10.1038/s41467-018-07038-8. ISSN 2041-1723. PMC 6218482. PMID 30397200.
- ↑ Anzai, Jun-Ichi; Osa, Tetsuo (1994). "Photosensitive artificial membranes based on azobenzene and spirobenzopyran derivatives". Tetrahedron. 50 (14): 4039–4070. doi:10.1016/S0040-4020(01)86704-1.
- ↑ Folgering, Joost H. A.; Kuiper, Johanna M.; de Vries, Alex H.; Engberts, Jan B. F. N.; Poolman, Bert (2004). "Lipid-Mediated Light Activation of a Mechanosensitive Channel of Large Conductance" (PDF). Langmuir. 20 (17): 6985–6987. doi:10.1021/la048942v. PMID 15301476.