刺甲龍屬
刺甲龍(屬名:Scolosaurus,意為「尖樁蜥蜴」)是甲龍科的一屬恐龍,生存於白堊紀晚期的拉臘米迪亞,化石在加拿大亞伯達省坎潘階中期(約7400萬年前)的恐龍公園組下部層位和老人組上部層位都有發現過。屬下包含兩個物種:模式種庫氏刺甲龍(Scolosaurus cutleri)、王座刺甲龍(Scolosaurus thronus)。有著複雜曲折的分類歷史,經常與近緣的包頭龍、倍甲龍混淆,其中過去包頭龍的重建其實大多是依照刺甲龍的形象。[2][1]
| 刺甲龍屬 化石时期:晚白堊世 | |
|---|---|
 | |
| 王座刺甲龍正模標本的骨架模型,展示於皇家蒂勒爾博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 科: | †甲龙科 Ankylosauridae |
| 亚科: | †甲龙亚科 Ankylosaurinae |
| 属: | †刺甲龍屬 Scolosaurus Nopcsa, 1928 |
| 模式種 | |
| †庫氏刺甲龍 Scolosaurus cutleri Nopcsa, 1928 | |
| 其他種 | |
| |
| 異名 | |
| |
發現及研究歷史

自1912年起,化石獵人兼商人威廉·庫勒(William Edmund Cutler)在加拿大亞伯達省展開挖掘活動。1914年他和當地牧人在死宅谷(Deadlodge Canyon)附近的80號採石場挖出了一具甲龍類骨骼。[2][3]當他試圖切割化石岩塊的基部時,突然翻倒壓碎了他的胸部,造成永久性創傷。[4]庫勒在幾天後獲救。而標本後來由查爾斯·斯騰伯格與其兒子們進行清復,最後被英國自然史博物館的地質學家法蘭西斯·貝瑟(Francis Arthur Bather)買下,於1915年運往倫敦,並保存於英國自然史博館至今。當初確切的挖掘地點再也無從得知。根據當時的報導,化石是從紅鹿河(Red Deer River)河谷樂傑渡口(Happy Jack Ferry)以南100英呎的懸崖面挖出來的,那裡現在屬於恐龍省立公園的一部分。2007年發現了最初的採石場標誌,但可能是已經被沖入峽谷深處的。木樁附近有個採石場,但與原始地圖上繪製的位置有所差異。在此地找尋其他可能的進一步材料仍無所獲。
1928年,弗朗茨·諾普札命名並敘述了模式種庫氏刺甲龍(Scolosaurus cutleri)。屬名源自古希臘語的σκῶλος/skoolos(刺),代表動物裝甲上長著刺。種名紀念發現者庫勒,[3]他已於1925年的非洲探險死於瘧疾。
正模標本NHMUK R5161確切的最初發現地已不可考。原始地圖標記指向恐龍公園組下部層位。但2007年定位的採石場屬於較古老的老人組。兩個地層年代都可追溯至坎潘階時期,但並不完全同時,因為彼此被不連續面分隔開來,大致落在7640萬年前左右。[5]因此刺甲龍的年代可能比地層年輕約幾十萬年、或更老。標本是一具異常完整的骨架,幾乎保存了全身骨骼,甚至還有罕見原位保存的皮內成骨和皮膚印痕,僅缺少尾部末端、右前後肢、頸椎前部、頭骨。標本僅部分清復完成,難以進行檢驗研究,並一直展示於博物館內的高玻璃櫃右側,管理人員暫無意願將之取出,因為當初耗費了相當大的力氣來設置成這副模樣。
自從被敘述以後,刺甲龍這個名稱一直缺乏關注、乏人問津。1971年,瓦特·庫姆斯(Walter Coombs)在他的博士論文質疑道刺甲龍與包頭龍幾乎完全一樣,可能是後者的次異名,他並將所有坎潘階的北美甲龍科歸類於該屬。[6]然而在2010年與2013年,甲龍類專家維多莉亞·阿布爾指出,單一物種不可能橫跨如此廣泛的時空,並開始將所有曾歸入包頭龍的材料劃分開來。於是刺甲龍屬的有效性再度被承認。發現於加拿大的正模標本曾是刺甲龍的唯一化石,但阿布爾另外歸入了兩個來自蒙大拿州雙麥迪遜組的化石。[7]該地層與恐龍公園組的年代吻合,[8]但都沒有比NHMUK R5161更為古老,因此後者代表了北美洲已知最古老的完整甲龍科化石。她同時反駁道剛於2013年新命名、根據相同的雙麥迪遜組材料而建立的石生龍屬是刺甲龍的異名。[7]但石生龍命名者保羅·潘卡司基(Paul Penkalski)和威廉·布洛斯(William Blows)雖然承認刺甲龍的有效性,卻不認同兩者是同一物種的觀點。[5]由於石生龍的關鍵性鑑定特徵位於頭骨,而刺甲龍沒有頭骨保存,造成爭議變得複雜化。歸入刺甲龍的蒙大拿標本包括MOR 433(石生龍的正模標本)、FPDM V-31、NSM PV 20381、TMP 2001.42.19以及USNM 7943。
2018年,潘卡司基將AMNH 5404標本,一個含有頭骨的骨骼歸入庫氏刺甲龍,並發表了一份與之前阿布爾截然不同的物種鑑定研究。同一年潘卡司基也命名了第二個種:王座刺甲龍(Scolosaurus thronus),種名取自拉丁語的throne(王位),代表其正模標本出土於恐龍公園組的上部層位,年代約7600萬年前,且發現地是一座很高的小山丘。王座刺甲龍的正模標本ROM 1930於1914年由喬治·斯騰伯格所發掘,後來長期歸類於包頭龍。他另外歸入了兩個標本:CMN 268(一塊頸部碎片)與TMP 1998.53.1(一塊有褶皺紋理的皮內成骨)。[1]
分類
1928年諾普薩將標本分類至甲龍科,並繪製了與倍甲龍的化石形態比較圖。[3]1971年庫姆斯表示坎潘階的北美洲只存在過單一個甲龍科物種,他將賴氏無齒甲龍、銳鱗倍甲龍、庫氏刺甲龍都歸入衛甲包頭龍,但未提供任何關於這些成為異名的理由。[6]庫氏刺甲龍是衛甲包頭龍的異名這個觀點被廣泛接受,於是NHMUK R.5161長期被歸在包頭龍。但潘卡司基和布洛斯2013年發表於加拿大地科學報對刺甲龍的重新描述論文承認該屬的有效性,他們得出結論為刺甲龍可透過頸部裝甲形態、其他裝甲細節、前肢結構上的差異來與包頭龍區別開來,同時指出刺甲龍與倍甲龍也在骨盆、裝甲上有所不同。[2]因為刺甲龍正模標本的完整度,自1971年以來已成為許多包頭龍重建形象的參考基礎,因此大部分流傳的包頭龍復原圖其實是刺甲龍的。
2013年阿布爾等人的研究發現石生龍無法與刺甲龍區別開來,因此將前者視為後者的疑名。[7]但這個觀點引發爭議,後來石生龍被認為是有效屬。[5]所以大部分被歸入刺甲龍屬的標本可能實際上是獨立的石生龍屬。


以下演化樹取自阿布爾和菲力·柯里2015年甲龍亞科系統發生學分析的綜合版本:[9]
| 甲龍亞科 Ankylosaurinae |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||


以下另一版本演化樹則為2017年阿布爾和大衛·伊凡斯(David Evans)的甲龍亞科支序分類學分析:[10]
| 甲龍亞科 Ankylosaurinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
归入材料

1874年,G·M·道森(G. M. Dawson)在阿尔伯塔省法国人组的牛奶河地区挖出标本USNM 7943。其收集于大约7060至6600万年前晚白垩世马斯特里赫特阶的陆地沉积物中。该标本由部分第一颈环(该恐龙颈部的一部分)组成。2013年,阿博尔和柯里进行一次详细的甲龙科系统发育分析,将该材料归入刺甲龙。[11]标本现保存于华盛顿特区的史密森学会。
1928年,乔治·斯腾伯格(George F. Sternberg)在蒙大拿州冰川县双麦迪逊组上层的蒙大拿神翼龙模式产地收集标本USNM 11892。[12]材料包含部分颅骨,收集于大约7400万年前坎帕阶期间沉积的海岸砂岩沉积物中,亦保存于史密森学会。
其他归入刺甲龙的标本包括FPDM V-31、NSM PV 20381及TMP 2001.42.9。FPDM V-31和TMP 2001.42.9均为颅骨,但保存状况不同。NSM PV 20381包含颅骨、背椎、尾椎、肋骨、两个肩胛骨、两个髂骨、部分坐骨及两个股骨、两个胫骨及腓骨。
鉴别特征
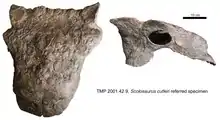

根据阿博尔和柯里(2013年)的研究,刺甲龙(包括双麦迪逊组的材料)可通过如下特征与其它甲龙亚科进行区分:
- 鳞状角成比例地更长、向后倾斜且有明显的顶端(与赖氏无齿甲龙和卫甲包头龙不同)
- 鳞状骨及方轭骨底部存在一个较小的圆形caputegula(与卫甲包头龙不同)
- 髂骨髋臼后突成比例地更长(与赖氏无齿甲龙、锐鳞倍甲龙和卫甲包头龙相比)
- 成比例较大、中部突起较低的圆形中段皮内成骨及颈半环上收缩的半月形近/远端皮内成骨(与赖氏无齿甲龙和卫甲包头龙不同)
- 骶肋指向侧面(与锐鳞倍甲龙不同)
- 皮内成骨呈圆锥形,顶端位于尾部前外侧的中间位置(与锐鳞倍甲龙不同)
- 尾槌突起在背视图中呈圆形,不像无齿甲龙这样宽大于长,也不像倍甲龙这样长大于宽
- 鼻孔指向前方及鳞状角和眶上骨之间没有连续的龙骨(与大面甲龙不同)
古生态学

栖息地
氩-氩放射定年显示双麦迪逊组沉积于8350至7060万年前晚白垩世坎帕阶期间,地理范围相当于现在的蒙大拿州西北部。[13]如果石生龙和刺甲龙是同种生物,那就意味着刺甲龙存在大约300万年。双麦迪逊组与阿尔伯塔西南部的贝利河组和东部的帕考基组相关。双麦迪逊组由西部内陆海道西海岸与科迪勒拉掩冲带东移边缘之间的河流及三角洲沉积而成。自中白垩世起,北美被这条海道一分为二,蒙大拿州和阿尔伯塔省的大部分地区皆在水面以下。然而落基山脉的隆起迫使海道向东并向南后退。河流自山上流下,携带沉积物汇入海道,形成双麦迪逊组和朱迪斯河组。大约7300万年前,海道开始向西并向北再次推进,整个地区均被熊爪海覆盖,形成如今纵贯美国西部和加拿大的熊爪页岩,该地层覆于双麦迪逊组之上。[14][15]地层下方是维吉尔砂岩的近岸沉积物。岩性、无脊椎动物群及植物和花粉数据显示,双麦迪逊组沉积于季节性半干旱气候中,可能位于科迪勒拉高地的雨影区。该地区拥有漫长旱季及温暖的气温。上双麦迪逊组的广阔红色岩层及钙质层是至少季节性干旱气候的证据。
古动物群

刺甲龙与其他恐龙共享古环境,如鸭嘴龙科的亚冠龙、始无冠龙、钩鼻龙、短脚龙、泥隐龙、原栉龙和慈母龙以及甲龙类的埃德蒙顿甲龙。[16]西部的火山喷发导致火山灰周期性覆盖此地区,造成大规模死亡,同时为未来的植物生长肥沃土壤。海平面波动还朱迪斯河群的不同时期及地区形成的各种其它环境,包括近海和近岸海洋栖息地、沿海湿地、三角洲和泻湖以及内陆漫滩。与其它两个地层相比,双麦迪逊组沉积在更远内陆区的更高海拔处。[17]该地区共存着各种各样的角龙类,包括河神龙、短角龙、赤角龙、野牛龙、倾角龙和刺丛龙。食肉动物包括一种未命名的伤齿龙科,可能是细爪龙、驰龙科的斑比盗龙、蜥鸟盗龙和大型暴龙科惧龙及蛇发女怪龙。[18]
双麦迪逊组和朱迪斯河群良好的脊椎动物化石记录是动物丰富、周期性自然灾害和大量沉积物沉积的综合结果。存在多种类型的淡水和淡咸水鱼,包括鲨鱼、鳐鱼、鲟鱼、鲈鱼等。该地区存在多种水生两栖动物和爬行动物遗骸,包括双壳类、腹足类、青蛙、蝾螈、海龟、鳄龙和鳄鱼。陆生蜥蜴包括叉舌蜥科、石龙子、巨蜥和蛇蜥科也被发现。像蒙大拿神翼龙和大鸟翼龙这样的翼龙以及像迷惑鸟和鸟龙鸟这样的鸟类在头顶上飞翔。几种哺乳动物如多瘤齿兽目的间异兽与恐龙在双麦迪逊组和构成朱迪斯河群的各种其它地层中共存。当地还发现一种驰龙科的蛋化石。水资源充足时该地区可养活大量动植物,但周期性干旱往往造成大规模死亡。[19]
参见
- 甲龙类研究历史
- 2018年古生物学研究
參考來源
- Paul Penkalski. . Neues Jahrbuch für Geologie und Paläontologie - Abhandlungen. 2018, 287 (3): 261–306. doi:10.1127/njgpa/2018/0717.
- Penkalski, P.; Blows, W. T. . Canadian Journal of Earth Sciences. 2013, 50 (2): 130110052638009. doi:10.1139/cjes-2012-0098.
- Nopcsa, B. F. . Geologica Hungarica, Series Palaeontologica. 1928, 1 (1): 1–84.
- Tanke, Darren H. . M.J. Ryan; B.J. Chinnery-Allgeier; D.A. Eberth (编). . Bloomington: Indiana University Press. 2010: 541–50 [2021-07-13]. ISBN 978-0-253-35358-0. (原始内容存档于2021-07-13).
- Penkalski, P. . Acta Palaeontologica Polonica. 2013. doi:10.4202/app.2012.0125.
- Coombs W. (1971) The Ankylosauridae. Ph.D. thesis, Columbia University, New York, NY, 487 p.
- Arbour, V. M.; Currie, P. J. Farke, Andrew A , 编. . PLoS ONE. 2013, 8 (5): e62421. PMC 3648582
 . PMID 23690940. doi:10.1371/journal.pone.0062421.
. PMID 23690940. doi:10.1371/journal.pone.0062421. - Rogers, R.R.; Swisher, III; Horner, J.R. . Canadian Journal of Earth Sciences. 1993, 30 (5): 1066–1075. doi:10.1139/e93-090.
- Arbour, V. M.; Currie, P. J. . Journal of Systematic Palaeontology. 2015, 14 (5): 1–60. doi:10.1080/14772019.2015.1059985.
- Arbour, Victoria M.; Evans, David C. . Royal Society Open Science. 2017, 4 (5): 161086. PMC 5451805 . PMID 28573004. doi:10.1098/rsos.161086.
- Cope, E. D. . Proceedings of the Academy of Natural Sciences of Philadelphia. 1875, 27: 2–3.
- K. Padian, A. J. de Ricqles, and J. R. Horner. 1995. Bone histology determines identification of a new fossil taxon of pterosaur (Reptilia: Archosauria). Comptes Rendus de l’Academie des Science, Serie II (320)77-84
- Eberth, David A. . Currie, Philip J.; Padian, Kevin (编). . San Diego: Academic Press. 1997: 199–204. ISBN 978-0-12-226810-6.
- English, Joseph M.; Johnston, Stephen T. (PDF). International Geology Review. 2004, 46 (9): 833–838 [2022-06-28]. Bibcode:2004IGRv...46..833E. S2CID 129901811. doi:10.2747/0020-6814.46.9.833. (原始内容 (PDF)存档于2011-06-07).
- Rogers, Raymond R. . Currie, Philip J.; Padian, Kevin (编). . San Diego: Academic Press. 1997: 199–204. ISBN 978-0-12-226810-6.
- Rogers, Raymond R. . PALAIOS. 1990, 5 (5): 394–413. JSTOR 3514834. doi:10.2307/3514834.
- Falcon-Lang, Howard J. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2003, 199 (3–4): 299–314. Bibcode:2003PPP...199..299F. doi:10.1016/S0031-0182(03)00539-X.
- "3.11 Montana, United States; 2. Lower Two Medicine Formation," in Weishampel, et al. (2004).
- Rogers, R. R. . PALAIOS. 1990, 5 (5): 394–41. Bibcode:1990Palai...5..394R. JSTOR 3514834. doi:10.2307/3514834.