栉龙属
櫛龍(屬名:Saurolophus,發音:/sɔːˈrɒləfəs/,意為「蜥蜴冠飾」,或譯龍櫛龍、蜥嵴龍、蜥冠龍)是種大型鴨嘴龍科恐龍,生存於晚白堊世的北美洲與亞洲,化石出土於加拿大的馬蹄鐵峽谷層與蒙古的耐梅蓋特層,年代約7000萬至6800萬年前。櫛龍是已知少數同時分布在多個大陸的恐龍物種之一。模式種奧氏櫛龍(S. osborni)於1912年由巴納姆·布朗根據加拿大的化石而命名。第二個有效種窄吻櫛龍(S. angustirostris)所知來自蒙古的數個標本,並由阿納托利·羅特傑斯特文斯基所描述。櫛龍最明顯的特徵是頭骨朝後上方45度角延伸的尖狀冠飾。櫛龍是相對大型的植食性動物,身長約8至13公尺,可用二足或四足行走。目前已發現多具完整的化石,因此能有較全面的了解,也因而成為鴨嘴龍科底下主要演化支之一櫛龍亞科的依據。
| 栉龙属 化石时期:晚白堊世 | |
|---|---|
 | |
| 窄吻櫛龍的骨架模型,展示於名古屋市科學館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 亚目: | †鸟脚亚目 Ornithopoda |
| 科: | †鴨嘴龍科 Hadrosauridae |
| 亚科: | †栉龙亚科 Saurolophinae |
| 族: | †栉龙族 Saurolophini |
| 属: | †栉龙属 Saurolophus Brown, 1912 |
| 模式種 | |
| †奧氏櫛龍 Saurolophus osborni Brown, 1912 | |
| 種 | |
| |
發現史


1911年巴納姆·布朗發現了首個被描述的櫛龍化石,是為編號AMNH 5220的正模標本,其保存了幾乎完整的骨骼部位,並成為加拿大第一次發現接近完整的恐龍骨骸;目前展示於美國自然歷史博物館。該標本出土於亞伯達省紅鹿河妥曼渡口(Tolman Ferry)附近的馬蹄峽谷層(當時舊稱為愛德蒙頓層/Edmonton Formation),地質年代屬於上白堊紀馬斯垂克階早期。布郎隨即花了些時間來研究他所找到的化石,[1][2]並為其建立了專屬的櫛龍亞科。[3]櫛龍在早期成為其他鴨嘴龍科的主要參考對象,例如原櫛龍(意即「櫛龍之前」)與副櫛龍(意即「接近櫛龍」)。然而櫛龍本身卻很少有額外化石被發現及描述。

反而後來在亞洲發現了豐富的化石來提供進一步資訊。最初的發現十分可疑,是來自中國黑龍江省的部份坐骨碎片,於1930年由里亞賓寧等人命名為克氏櫛龍(S. kryschtofovici)。[4]不過此後不久,於1946至1949年蘇聯-蒙古聯合古生物考察活動期間,便在蒙古同樣屬於馬斯垂克階早期的耐梅蓋特層挖出保存較好的大型骨骼,即成為1952年由羅特傑斯特文斯所描述的窄吻櫛龍(S. angustirostris)。[5]此外亦找到其他屬於不同生長階段的相關化石,使窄吻櫛龍成為亞洲已知最豐富的鴨嘴龍科之一。[6]
物種

目前有兩個有效種:模式種奧氏櫛龍(S. osborni)、窄吻櫛龍(S. angustirostris)。奧氏種由巴納姆·布郎於1912年命名,已知標本包括一個頭骨與骨骼、另外兩個完整的頭骨、一些頭骨碎片。窄吻櫛龍由羅特傑斯特文斯基於1952年命名,已知有至少15個標本。[7]後者與前者的差異在於頭骨的一些細部特徵、以及皮膚化石上的鱗片排列模式。窄吻種的頭骨較長(長於奧氏種的20%)、吻部前端的前上頜骨角度更為朝上。[8]窄吻種延背部至尾部中線有一排明顯的矩形鱗片,這在奧氏種中並未保存下來。窄吻種尾部側面鱗片呈垂直排列,生前可能對應著有顏色的條紋。奧氏種的背部鱗片則較為光滑,尾側鱗片呈輻射狀排列,生前可能對應著斑點或斑駁狀的顏色。[9]
克氏櫛龍(S, kryschtofovici)由阿納托利·里亞賓寧於1930年命名,目前不被承認為有效種,而是疑名[10][7]或窄吻種的異名[6](雖然前者較早命名)。[11]
窄吻種長期以來沒有經過充分的描述,像是區分出與奧氏種差異的自衍徵、獨有特徵等,直到2011年貝爾(Phil R. Bell)對該物種進行了重新檢視。他在同一年稍早發表的文獻中提到,在1952年羅特傑斯特文斯基以及1981年奧斯穆斯卡和馬瑞沿斯卡的研究都沒有提供足夠全面的描述來比較兩物種的差異。[12]
1939至1940年期間在加州馬斯垂克階晚期的莫瑞諾層出土兩具部分骨骼,當時標名為「cf. Saurolophus sp.」(cf.是拉丁文conformis的縮寫,意指其為櫛龍的近似種)。2010年其中一個頭骨被歸到愛德蒙托龍。[13]2013年的研究將兩個標本建立為新物種莫氏櫛龍(S. morrisi)。[11]2014年莫氏種被建立成獨立的奧氏櫛龍屬(Augustynolophus)。[14]
描述
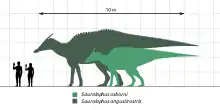


已知櫛龍的遺骸涵蓋了近乎完整的全身骨骼,使研究人員能清晰的認識其身體結構。較罕見的亞伯達奧氏種身長約8.2公尺,頭骨長1公尺,[15][16]估計體重約3噸。[16]蒙古窄吻種體型較大,身長達到13公尺,發現的化石也較豐富。估計體重可能高達11噸。[16]已知最大的窄吻種頭骨測量長1.22公尺。[12]除了明顯的體型差異,兩個物種基本上非常相似,但缺乏充分研究導致兩者難以區分。[10]
頭骨



櫛龍最特別的地方是牠的頭冠,長而尖狀,從眼睛上方開始,往頭後上方45度角傾斜伸出。頭冠通常被描述成實心,但僅限於頂端,其餘內部可能具有呼吸或調節體溫功能的腔室。[17]頭冠絕大部分由鼻骨構成,窄吻種的鼻骨為實心。年輕個體的頭冠較小。成體頭冠的橫切面為圓潤的三角形,向後突出超越頭骨邊緣,額骨與前額骨的細長突起延頭冠下側延伸,可能用來加固。頭冠末端有鼻骨膨脹。[12]
窄吻種的正模標本包含頭骨與顱後骨骼,因此其頭骨得到很充分的描述。貝爾等人(2011)重新檢驗了該物種,發現頭骨在櫛龍亞科為典型,且比任何奧氏櫛龍的頭骨都大得多。櫛龍亞科異常的特徵在於從頭蓋骨斜後向上延伸出長而突出的實心頭冠,不同於賴氏龍亞科,櫛龍的頭冠完全由鼻骨構成。前上頜骨幾乎占據整個頭骨長度的一半,兩側佈滿小孔洞。前上頜骨前方的接觸位置僅在成體是癒合的。鼻骨長於前上頜骨,且是頭骨中最長的一部分。它們一同構成整條頭冠,且彼此從未癒合在一塊。[12]
分類
描述模式種的布朗將櫛龍置於糙齒龍科(Trachodontidae)/鴨嘴龍科底下的專屬演化支櫛龍亞科。當時其中還包含後來成為賴氏龍亞科代表的冠龍和亞冠龍。[3]布朗認為櫛龍臀部坐骨有個尖狀擴張,如同現今定義賴氏龍亞科的特徵,但後來發現這可能是將其他賴氏龍亞科的坐骨給混淆的結果。此外他還誤認為櫛龍與賴氏龍亞科的頭冠都是由相同骨頭構成。[18]
在2010年以前,大部分文獻都將櫛龍歸入鴨嘴龍亞科,即所謂「頭頂平坦的鴨嘴龍」。2010年櫛龍亞科開始被重新採用,因為發現鴨嘴龍屬本身在鴨嘴龍科的分化時間,早於傳統定義的鴨嘴龍亞科與賴氏龍亞科的分化時間,於是鴨嘴龍亞科僅限於鴨嘴龍本身的分支,而其餘傳統上的鴨嘴龍亞科物種則被改歸入櫛龍亞科,因為櫛龍是其中最早命名的物種,足以成為演化支的依據。後來新的櫛龍亞科定義為:具有與櫛龍類似的骨盆、以及(大部分為)實心的頭冠。[19]
以下櫛龍亞科支序圖取自皮耶妥·馬奎茲(Alberto Prieto-Márquez)等人2013年的研究:[20]
| 櫛龍亞科 Saurolophinae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生物學
頭冠功能

關於櫛龍特殊的尖狀頭冠已提出多種功能的解釋。布朗將其比作變色龍的頭冠,並成為一道肌肉附著的區域,可能與某種背部皮膜構成一片連接面,類似雙脊冠蜥的情況。彼得·多德森認為這與其他有頭冠的鴨嘴龍科一樣用於性別展示。[21]馬瑞沿斯卡和奧斯穆斯卡指出頭罐底部空心,顯示構成面積擴張的呼吸腔室,並有助於體溫調節。[17]詹姆斯·霍普森支持視覺展示的論點,並進一步表示鼻孔上的中空皮瓣可能作為共鳴腔或加強視覺功能的可能性。[22]此理論已受到大眾恐龍作品所採納,如大衛·諾曼詳細探討了鴨嘴龍科的展示功能,並繪製相關復原圖加強說明。[23]
個體發育





雖然過去對櫛龍的成長速率所知甚少,最近在戈壁沙漠一處稱為「龍之墓」的遺址發現了櫛龍群聚築巢的化石。[24]2015年雷納·德威爾(Leonard Dewaele)在科學期刊《PLOS ONE》發表了關於MPC-D 100/764標本的描述——一個小型部分巢穴,當中含有幾隻窄吻櫛龍幼體。研究者識別出至少三到四隻幼體以及兩個蛋殼碎片,由這些個體頭骨尺寸不到成體的5%判斷死亡時尚處於剛孵化、住在巢穴的早期發育階段。德威爾等人解釋了窄吻櫛龍的個體發育過程,成體明顯的頭冠在幼體時幾乎還沒發育出現,隨成長吻部將按比例增長、眼眶變得更偏橢圓形、額骨隆起日益隱沒、頭冠突起抬升。[25]
社會行為

2018年貝爾等人描述了位於奧譚烏爾二區(Altan Uul II)耐梅蓋特層的知名龍之墓遺址,當中有個大型窄吻櫛龍屍骨層,由單一物種獨佔,存在三種不同尺寸級別(幼體、亞成體、成體),至少可辨識出達21隻個體。屍骨層涵蓋面積最少有2000平方公尺,代表可能有超過100隻櫛龍參與該埋藏事件。他們討論道該屍骨層反映一場群體性的大規模死亡事件,並提供了櫛龍是群居動物的首個證據,至於族群死亡的確切條件及原因則無法得知。貝爾等人還提到窄吻種已經有在龍之墓的群居證據,奧氏種卻還沒發現,不過群居行為顯然在鴨嘴龍科是普遍情況。[25]
古病理學
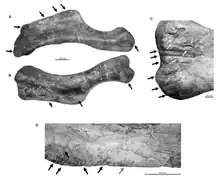
2011年洪大衛(David W.E. Hone)和渡部真人記載了來自耐梅蓋特布津察夫(Bügiin Tsav)一具近乎完整的櫛龍骨骼(MPC-D 100/764)的左肱骨,該處被同地的特暴龍咬到損毀。由於骨骼其餘部位均未受到破壞,判斷特暴龍可能是在啃食櫛龍屍體。像特暴龍這類大型掠食者不太可能在有整個軀體的情況下只在肱骨留下稀疏的咬痕,且肱骨處的咬痕顯示三種不同進食方式:穿刺、拖曳、撕扯。洪和渡部指出咬痕主要位於三角肌嵴,反映那隻特暴龍曾採取積極作為來選擇用何種方式來啃食屍體。[26]
食性
作為一種鴨嘴龍科,櫛龍是植食性動物,以各種植物為主食,可用二足也可四足行走。其頭骨能作出類似咀嚼功能的磨削動作,植物會先被寬闊的嘴喙切割,並由一道類似臉頰的構造夾在口內。具有持續汰換的數百顆牙齒所構成的齒系,其中僅有相對少數的牙齒是持續在使用。取食範圍從地面延伸到上方約4公尺處。[7]
古生態學
馬蹄鐵峽谷
奧氏種來自馬蹄鐵峽谷層上部第四層位。當時受到西部內陸海道的影響,顯示出明顯的海洋性氣候。[28]奧氏種可能偏好棲息於內陸環境。[7]與之共存的恐龍包括:鳥腳類的高棘亞冠龍、瓦氏帕克氏龍;甲龍科的賴氏無齒甲龍;厚頭龍科的古氏圓頭龍;似鳥龍科的短背似鴯鶓龍以及似鴕龍未命名種;小型獸腳類有馬氏野蠻盜龍、北方亞伯達爪龍;暴龍科的肉食亞伯達龍。[29]以上恐龍種群構成了愛德蒙頓陸地脊椎動物群的一部分。[28]一份2001年的研究指出,奧氏櫛龍與華麗準角龍屬於內陸種群的關聯代表;與之對應的沿海種群代表則是加拿大厚鼻龍與皇家愛德蒙托龍。[30]然而後來發現櫛龍與準角龍之間的關聯性不太可靠,因為準角龍僅產自馬蹄鐵峽谷層較下部層位,當時西部內海的卓姆赫勒大海侵(Drumheller Marine Tongue)還沒發生。[31]
耐梅蓋特

窄吻櫛龍是耐梅蓋特層最大型的植食性動物之一,當地缺乏大型角龍科,但有蜥腳類及更多樣化的獸腳類。不像其他蒙古地層(如產出知名伶盜龍與原角龍的德加多克塔層),耐梅蓋特層是水源充沛的環境,類似亞伯達省的恐龍公園層。[28]對該地層進行檢驗,發現其岩相表明存在溪流、河道、泥灘、淺湖,沉積物並顯示出豐富的棲地類型,提供充足且多樣化的食物來源,足以孕育大量恐龍種群的生存。[32]窄吻櫛龍與以下恐龍共享棲地:較罕見的鴨嘴龍科巴思缽氏龍;頭部平坦的厚頭龍科平頭龍和傾頭龍;大型甲龍科美甲龍;稀有的泰坦巨龍類耐梅蓋特龍與後凹尾龍;阿瓦拉慈龍科的單爪龍;三種傷齒龍科包含扎納巴扎爾龍;數種偷蛋龍類如瑞欽龍耐梅蓋特母龍;似鳥龍科的似鵝龍、似雞龍;巨大的獸腳類有恐手龍、鐮刀龍、暴龍科的特暴龍。[29]
窄吻櫛龍在耐梅蓋特層相當常見,很可能是當地優勢的大型植食性動物;相反地,馬蹄鐵峽谷層的奧氏櫛龍則相對罕見,且很可能受到其他鴨嘴龍科(如亞冠龍)的競爭。
延伸閱讀
- 鴨嘴龍類研究歷史
參考來源
- Brown, Barnum. . Bulletin of the American Museum of Natural History. 1912, 31 (14): 131–136. hdl:2246/1401.
- Brown, Barnum. . Bulletin of the American Museum of Natural History. 1913, 32 (19): 387–393. hdl:2246/1415.
- Brown, Barnum. . Bulletin of the American Museum of Natural History. 1914, 33 (55): 559–564. hdl:2246/1734.
- Riabinin, Anatoly Nikolaenvich, N. . Mémoir, Société Mineral Russia. 1930, 59: 41–51 (俄语).
- Rozhdestvensky, Anatoly K. [A new representative of the duck-billed dinosaurs from the Upper Cretaceous deposits of Mongolia]. Doklady Akademii Nauk SSSR. 1952, 86 (2): 405–408 (俄语).
- Glut, Donald F. . . Jefferson, North Carolina: McFarland & Co. 1997: 788–789. ISBN 0-89950-917-7.
- Horner, John R.; Weishampel, David B.; Forster, Catherine A. . Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (编). 2nd. Berkeley: University of California Press. 2004: 438–463. ISBN 0-520-24209-2.
- Bell, Phil R. (PDF). Acta Palaeontologica Polonica. 2011, 56 (4): 703–722. doi:10.4202/app.2010.0061
 .
. - Bell, P.R. . PLOS ONE. 2012, 7 (2): e31295. Bibcode:2012PLoSO...731295B. PMC 3272031 . PMID 22319623. doi:10.1371/journal.pone.0031295 .
- Norman, David B.; Sues, Hans-Dieter. . Benton, Michael J.; Shishkin, Mikhail A.; Unwin, David M.; Kurochkin, Evgenii N. (编). . Cambridge: Cambridge University Press. 2000: 462–479. ISBN 0-521-55476-4.
- Albert Prieto-Márquez; Jonathan R. Wagner. . Acta Palaeontologica Polonica. 2013, 58 (2): 255–268. doi:10.4202/app.2011.0049.
- Bell, P. R. . Acta Palaeontologica Polonica. 2011, 56 (4): 703–722. doi:10.4202/app.2010.0061 .
- Bell, P.R.; Evans, D.C. . Canadian Journal of Earth Sciences. 2010, 47 (11): 1417–1426. Bibcode:2010CaJES..47.1417S. doi:10.1139/E10-062.
- Prieto-Márquez, Albert; Wagner, Jonathan R.; Bell, Phil R.; Chiappe, Luis M. . Geological Magazine. 2014, 152 (2): 225–241. S2CID 131049979. doi:10.1017/S0016756814000284.
- Lull, Richard Swann; Wright, Nelda E. . Geological Society of America Special Paper 40. Geological Society of America. 1942: 226.
- Paul, Greg. . New Jersey: Princeton University Press. 2010: 335.
- Maryańska, Teresa; Osmólska, Halszka. (PDF). Palaeontologia Polonica. 1981, 42: 5–24 [2022-03-14]. (原始内容 (PDF)存档于2021-09-26).
- Sternberg, Charles M. . Journal of Paleontology. 1954, 28 (3): 382–383.
- Prieto-Márquez, Alberto. . Zoological Journal of the Linnean Society. 2010, 159 (2): 435–502. doi:10.1111/j.1096-3642.2009.00617.x .
- Prieto-Márquez, A.; Wagner, J.R. . Acta Palaeontologica Polonica. 2013, 58 (2): 255–268. doi:10.4202/app.2011.0049 .
- Dodson, Peter. . Systematic Zoology. 1975, 24 (1): 37–54. JSTOR 2412696. doi:10.2307/2412696.
- Hopson, James A. . Paleobiology. 1975, 1 (1): 21–43. doi:10.1017/S0094837300002165.
- Norman, David B. . . New York: Crescent Books. 1985: 122–127. ISBN 0-517-46890-5.
- (新闻稿). PLOS. October 14, 2015 [2022-03-14]. (原始内容存档于2022-03-14) –Science Daily.
- Dewaele, Leonard; Tsogtbaatar, Khishigjav; Barsbold, Rinchen; Garcia, Géraldine; Stein, Koen; Escuillie, François; Godefroit, Pascal. . PLOS ONE. October 14, 2015, 10 (10): e0138806. Bibcode:2015PLoSO..1038806D. PMC 4605499 . PMID 26466354. doi:10.1371/journal.pone.0138806 .
- Hone, D. W. E.; Watabe, M. (PDF). Acta Palaeontologica Polonica. 2011, 55 (4): 627−634 [2022-09-17]. doi:10.4202/app.2009.0133 . (原始内容存档 (PDF)于2021-09-19).
- Schmitz, L.; Motani, R. . Science. 2011, 332 (6030): 705–8. Bibcode:2011Sci...332..705S. PMID 21493820. S2CID 33253407. doi:10.1126/science.1200043.
- Dodson, Peter. . Princeton: Princeton University Press. 1996: 14–15. ISBN 0-691-05900-4.
- Weishampel, David B.; Barrett, Paul M.; Coria, Rodolfo A.; Le Loueff, Jean; Xu Xing; Zhao Xijin; Sahni, Ashok; Gomani, Elizabeth M.P.; Noto, Christopher N. . Weishampel, David B.; Dodson, Peter; Osmólska, Halszka (编). 2nd. Berkeley: University of California Press. 2004: 517–606. ISBN 0-520-24209-2.
- Lehman, T. M., 2001, Late Cretaceous dinosaur provinciality: In: Mesozoic Vertebrate Life, edited by Tanke, D. H., and Carpenter, K., Indiana University Press, pp. 310-328.
- Sullivan, R.M. and Lucas, S. G. (2006). "The Kirtlandian land-vertebrate "age"–faunal composition, temporal position and biostratigraphic correlation in the nonmarine Upper Cretaceous of western North America." Pp. 7-29 in Lucas, S. G. and Sullivan, R.M. (eds.), Late Cretaceous vertebrates from the Western Interior. New Mexico Museum of Natural History and Science Bulletin 35.
- Novacek, M. (1996). Dinosaurs of the Flaming Cliffs. Bantam Doubleday Dell Publishing Group Inc. New York, New York. ISBN 978-0-385-47775-8