棘刺龍屬
棘刺龍(屬名:Spinophorosaurus)是蜥腳下目的一屬恐龍,生存於侏羅紀中期,地理範圍相當於現今的尼日。首兩具標本是於2000年代由德國和西班牙團隊在艱辛的環境下挖掘出土。骨骼被帶往歐洲,並進行數位複製,使棘刺龍成為第一個使用3D列印技術製作出的骨架模型,未來將返還給尼日當局。兩件標本一共涵蓋了本物種骨骼的大多數部位,為其年代和地區已知最完整的基群蜥腳類之一。第一具標本於2009年作為建立新屬新種尼日棘刺龍(Spinophorosaurus nigerensis)的正模標本;屬名意為「有棘刺的蜥蜴」,指一開始被認為尾部長有棘刺狀皮內成骨;種名「來自尼日的」即指發現地。來自同地區的一個少年蜥腳類個體後來也被歸入本屬。
| 棘刺龙属 | |
|---|---|
 | |
| 3D列印製作出的棘刺龍正模標本骨架模型,展示於布倫斯維克國家自然史博物館;地板上的化石則為真品 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | †蜥脚形亚目 Sauropodomorpha |
| 下目: | †蜥腳下目 Sauropoda |
| 演化支: | †沉重龙类 Gravisauria |
| 属: | †棘刺龙属 Spinophorosaurus Remes et al., 2009 |
| 模式種 | |
| †尼日棘刺龍 Spinophorosaurus nigerensis Remes et al., 2009 | |
亞成體的正模標本估計身長13公尺,而副模標本長約14公尺。這些個體的肩高估計可達到4公尺,體重約7噸。腦殼短、深而寬闊,神經解剖學某些方面界於基礎蜥腳形亞目和進階新蜥腳類之間的中等程度。牙齒呈勺狀/湯匙狀,齒冠頂部有大間隔的鋸齒狀,為蜥腳類的祖徵。棘刺龍的頸部是已知蜥腳類中最完整的之一,有13節頸椎。背部脊椎有許多小的中空內部腔室,是較晚期進階蜥腳類的典型特徵。尾部由強壯的肌肉組織作為動力,後部區域因人字骨較長且重疊而變得相當僵硬。一開始被認為是棘刺的骨頭,後來發現其實屬於鎖骨。
棘刺龍曾被分類為非常基礎的蜥腳類,或是屬於較進階的真蜥腳類。標本的解剖結構、年代和地點均指出,蜥腳類演化歷程的重大發展可能發生於北非,或許受到氣候區位以及植物生物地理學條件的影響。前庭系統特徵顯示,視覺、眼部協調、頭部、頸部活動力在棘刺龍身上非常重要。透過3D骨架模型來檢驗其運動程度。其中一項研究表示牠可能是高處攝植者,另一項研究則檢驗了可能的交配姿勢。棘刺龍軀幹前部脊椎神經弓和椎體之間的縫線較為複雜,是由於此區域承受的壓力可能最大。棘刺龍來自埃爾哈澤層,是個被認為屬於侏羅紀中期的地層,由大河谷系統中的河流和湖泊沉積物所構成。
發現歷史
尼日盛產豐富的恐龍群系,在經歷1960和70年代法國和義大利的考察活動後,逐漸成為科學界的矚目焦點。當中的重大發現包含敘述了下白堊紀禽龍類新屬豪勇龍。美國古生物學家保羅·塞里諾探勘了一個較早期的連續性岩層條拉輪組,於1999年至2003年間展開大規模的挖掘活動。最初以為該地層屬於下白堊紀,現在被證實為更古老的侏羅紀中期。塞里諾命名的新恐龍包含蜥腳類的約巴龍和獸腳類的非洲獵龍,兩者均來自條拉輪組;大多數發現都是在阿加德茲地區南部的提奎狄特懸崖(Falaise de Tiguidit)上找到的。塞里諾在馬連德將約巴龍的部分骨骼留在野外原址當作觀光景點。[2]:11–13[1]自2003年起,PALDES計畫(Paleontología y Desarrollo/Palaeontology and development/古生物學與發展)在阿加德茲地區南部展開挖掘活動。這項計畫是西班牙科學和人道主義機構間的合作,旨在將古生物學研究與當地發展計畫互相結合,如改善基礎建設、教育結構以及發展觀光旅遊產業,其中包括規劃在塔蒂賓(Tadibene)建設新的古生物博物館。[3]
2005年初,德國探險家烏里克·酋格(Ulrich Joger)和艾格·桑默(Edgar Sommer)在接獲圖阿雷格人關於在阿加德茲南部發現大型骨骼的通報後,前往當地的半沙漠地區進行探勘。桑默是救濟組織CARGO的創辦人,致力改善圖阿雷格人的教育體系;酋格則是生物學家兼布倫斯維克國家自然史博物館館長。他們在回程路上與一群圖阿雷格人聊天,後者便將前者指引至附近山區,在那裡到處散布著小骨頭碎片。此地區過去未曾發掘過恐龍,位於法萊斯·提奎狄特以北約30公里處,鄰近阿加德茲的阿德比希納鎮,屬於位於條拉輪組之下(故年代稍老些)的埃爾哈澤層的一部分。在經歷了1小時的搜索,酋格終於發現一個圓潤的骨頭尖端從地表突出,隨後經過進一步的挖掘,發現是一根完整的股骨,屬於後來即將發現的棘刺龍正模標本的一部分。不久之後便發現了關聯的肩胛骨和脊椎。當地的沉積物是堅硬但易碎的粉砂岩,可透過輕錘擊從骨頭上移除。[2]:17–27, 38[1][4]

接著酋格和桑默雇用當地的圖阿雷格人來支援,經歷兩天的搜索,發現了標本的大部分,包括一個近乎完整、關節仍連接的脊柱、數個肢骨以及骨盆。脊柱彎曲構成一個幾乎完整的圓形,尾部尖端處預計是頭骨的位置,但沒有找到頭骨。因為設備及挖掘許可證的缺乏,他們利用碎石將標本覆蓋起來作為防護,然後返回德國,計畫由布倫斯維克博物館展開全面的科學挖掘活動。2006年,尼日當局向博物館發出正式挖掘許可證,而博物館在因吉塔尼(Injitane)聚落位當地圖阿雷格兒童設立一所新學校,以作回報。2006年秋,桑默和酋格協同其他博物館同事,重返挖掘地點作準備,將骨盆其中一塊放入石膏,來測試設備與方法。在距離挖掘地點約1公里處,團隊還發現了獸腳類足跡。於2007年初找到了學校兼挖掘活動的贊助商。官方計畫稱為「Projekt Dino」,於2007年3月1日展開,當時兩輛裝載設備的卡車從布倫斯維克出發前往尼日,路線沿途經過西班牙、摩洛哥、茅利塔尼亞、馬利等國家,而直接穿越撒哈拉的較短路徑因恐怖攻擊的風險而作罷。團隊其餘部分由十名常設成員組成,搭乘飛機抵達。[2]:17–27, 38[1][5][6]這是歷經一世紀以來,德國首次的非洲恐龍考察活動。[1]:29[7]
與此同時,西班牙由埃爾切古生物博物館(Palaeontological Museum of Elche)領導的PALDES計畫團隊正在當地進行挖掘工作。2007年初,阿德比希納鎮長穆罕默德·艾奇卡(Mohamed Echika)允許PALDES團隊挖掘先前由德國人發現的骨骼;於是骨骼在其後被船運往西班牙。而德國團隊在毫不知情的狀況下,3月16日先鋒抵達時發現了一個空的挖掘現場(並呈現出專業挖掘的痕跡);卡車則於3月20日到達。雖然感到失望,德國團隊於3月17日,在與正模標本相距15公尺位置,發現了第二具棘刺龍標本,即後來的副模標本。一道可探溝渠區域內散佈著小骨頭碎片,很快就露出頜部和牙齒碎片;到第二天已經可以識別出肋骨、脊椎、肱骨、肩胛骨。3月19日八位當地挖掘助手加入團隊。3月20日卡車抵達前夕,已經耗盡最初200公升的儲備淡水,因為當地助手前一天晚上用水來沖洗,導致團隊成員暈倒。挖掘工作通常在中午12:00到15:00之間,溫度高達攝氏43至45度期間中斷。3月25日,除兩名德國成員外,其餘全部人都生病了,為腹瀉與血液循環問題所苦。在整個挖掘活動期間,透過照片和田野筆記來記錄進展狀況。[2]:29–45[8]

截至3月25日,將來成為副模標本的肱骨、肩胛骨、大部分肋骨都已經取出並由石膏包覆。雖然骨骼之下已經沒有進一步的骨頭可見,團隊仍清除60至80公分厚的沉積岩以確保所有化石都有收集到。[2]:60挖掘工作於4月2日完成,4月3日將化石打包好運往柯多努港。在同日艾奇卡向團隊透露第一批骨骼是在他的允許下由西班牙人挖走;並承諾將會指引團隊到阿加德茲以南80公里處提奎狄特懸崖的另一處化石遺址作為補償。團隊在那裡發現了可能的約巴龍後部骨架,但被迫遺留下最大一塊直到下一季再回來。為了防止其他人採集該岩塊,製作了一個爆炸性假人附著於化石之上,並附上西班牙語的警告標示。[2]:68–73[9]德國團隊於2008年的下一季重返並取回了岩塊;而西班牙PALDES團隊因圖阿雷格叛亂事件的爆發取消了當年的挖掘計畫。[2]:100, 107
兩具棘刺龍標本暫時保存於西班牙和德國的博物館。根據與尼日簽訂的合約,標本未來會交還給尼日,由尼亞美國家自然歷史博物館(Musée National Boubou Hama)與另一間較小的新建地方博物館所管理。[2]:143未來的副模標本於2007年3月18日抵達德國,布倫斯維克博物館承租了一座獨立工廠大樓以為了清修化石,費時兩年半。同一時間,德國與西班牙團隊已展開合作,一同準備一份聯合報告。德國團隊使用雷射掃描將兩件標本清修完成的骨骼和碎片給3D數位化。布倫斯維克博物館的標本僅70%完整,於是利用西班牙標本來填補缺失部位;過程中,人們發現西班牙標本是德國人最一開始發現並遺失的骨骼。3D掃描經過數位修復和還原變形後,以3D列印出來並組裝成骨架,於布倫斯維克博物館展出,成為透過3D列印製作出的第一個蜥腳恐龍骨架模型。在博物館主入口大門擺設了一個實體大小的棘刺龍復原模型,綽號為「Namu」(以博物館名稱為名)。[2]:79–85[10][11]西班牙團隊對正模標本的照片使用攝影測量法(從不同角度拍攝物件並拼湊起來)來合成個別的3D模型;[12]2018年艾爾切博物館展出一個尾椎。[13]2018年的論文摘要中,加席亞-馬汀尼茲(García-Martínez)和同事宣布他們透過保存較佳的第一、第五背椎來重建保存較差的第二背椎外形;這是依照形態測量學來完成的,從每個脊椎收集對應3D座標並進行統計分析。[14]

第一具骨骼(於兩間博物館分別編號:艾爾切為GCP-CV-4229、布倫斯維克為NMB-1699-R[15]),由德國古生物學家克里斯蒂安·雷梅斯(Kristian Remes)和其德國與西班牙研究團隊的同事,於2009年選定成為新屬新種尼日棘刺龍(Spinophorosaurus nigerensis)的正模標本。屬名由拉丁語的spina(尖刺)及希臘語的phoro(長著)加上sauros(蜥蜴)組成,全義即為「長著尖刺的蜥蜴」,名字由來為一開始被解釋為尾部長有帶棘刺的皮內成骨(但後來被認為屬於鎖骨)。種名意指尼日,即標本發現地。[16][1][17][18]正模標本包含:腦殼、眶後骨、鱗狀骨、方骨、翼骨、上隅骨、以及幾乎完整的顱後骨骼,僅缺乏胸骨、前臂、手部、指爪。第二具標本/副模標本NMB-1698-R由部分頭骨與不完整的顱後骨骼組成。副模標本有保存、而正模標本缺乏的部位包括:前上頜骨、上頜骨、淚骨、齒骨、隅骨、右側背椎、肱骨、腳趾。兩具標本因為骨骼重複部位相同,加上在同地層內骨骼彼此接近,被認為屬於同一分類單元。[1]在敘述發表當時,棘刺龍是北非最完整的侏儸紀中期蜥腳類,[1]且截至2018年都被認為是最完整的早期(非新蜥腳類)蜥腳類。[19]
2012年,PALDES團隊的艾卓安·帕拉莫(Adrián Páramo)和法蘭西斯科·歐特加(Francisco Ortega)報導了在距離兩具棘刺龍標本幾公尺處地面發現的一個小型蜥腳類骨骼(編號GCP-CV-BB-15標本);所有化石可能都來自同一地層。小型骨骼由14節脊椎(有些關節相連)組成,包括所有的頸椎及部分背椎。其椎體比棘刺龍的小20%,而神經椎體縫線是開放的,代表是隻幼體。小骨骼與棘刺龍有數項共同特徵,雖然有些在棘刺龍上找不到,但可以推測小骨骼屬於棘刺龍的幼年個體(特徵差異可能藉由個體發生學變化來解釋)。[20][15]
敘述

正模標本沿脊柱測量身長約13公尺,副模標本體型則大了約13%,測量約14公尺。[1][2]正模標本骨架的3D攝影測量模型從頭到尾長約11.7公尺,比例上與根據2D骨骼重建估計值有所差異。[12]這些個體達到的肩高估計值為4公尺,[2]體重約7噸。[1][21]正模標本頭骨的顱內和神經中樞縫線尚未癒合,代表是隻亞成體,而副模標本有完全癒合的神經中樞縫線。[1]
2020年,Vidal和同事根據數位骨架的設置來修正棘刺龍的姿勢。2009年的初始骨骼重建將恐龍呈現為水平姿勢,但數位重建則呈現更垂直的姿勢,即肩膀高、頸部抬升。[22]
頭骨

頭蓋骨的額骨中線癒合,不像頭骨其他縫線的狀況。在額骨、頂骨之間的縫線前約10毫米處,有個小的顱頂眼。頭骨後緣有個開放的後頂骨凹口,只有叉龍科、文雅龍有相同特徵。連接的額骨前部中線之後有個凹槽,代表從兩邊鼻骨到額骨之間可能有道叉狀延伸,尼日龍可能也有相同狀況,但這在一般蜥腳類中相當罕見。上顳孔(頭蓋骨一對主孔洞)寬度為長度的四倍;這類寬闊的孔是蜀龍和其他更衍化蜥腳類的典型特徵。頭骨後部的枕髁側凹,如同蜀龍。基結節(顱基底側一對用來附著肌肉的延伸)加大並朝側向,在蜥腳類中相當獨特。方骨後側沒有凹陷,是蜥腳類一項原始特徵,僅在塔鄒達龍發現過。匙狀的牙齒的特別之處在於,沿齒冠有著加大間隔的鋸齒狀,其中齒冠前緣鋸齒數目更多。[1][23]

棘刺龍腦殼前後寬闊而短,深度適中,整體尺寸相對較大。腦殼與其他侏儸紀蜥腳類有很大的差異,可能亞特拉斯龍除外。兩者的共同點為基蝶骨的基翼突大幅朝向後。一個正模標本腦腔的CT內模顯示出,與其他多數蜥腳類的相似處在於,橋腦和神經管彎曲(cerebral flexures)發育良好,腦下腺窩(pituitary fossa)大而長,以及相對厚的腦膜、硬腦膜靜脈竇儲存空間佔據了腦部結構。內耳骨迷路以長而纖細的半規管為特徵,與大椎龍、長頸巨龍相似。其神經解剖學大致介於基礎蜥腳形亞目和較進階的新蜥腳類之間。[23]
脊椎和肋骨

脊椎幾乎完整保存下來,且正模標本是少數包含完整頸部的蜥腳類。[24]頸部由13節頸椎構成。軀幹有12節胸椎和4節薦椎。尾部由超過37節尾椎構成。[1]複合材料、個別脊椎由下部椎體和上部神經弓構成。神經弓重要特徵包括上凸的神經棘(棘突)和側凸的橫關節突(diapophyses),兩者使脊椎前後視角呈T字形。將相鄰脊椎相連的關節突,前方為前關節突(prezygapophyses),後方為後關節突(postzygapophyses)。[25]
頸椎與約巴龍、鯨龍相似。牠們的椎體長度為寬度的3.1倍;與蜥腳類平均值相比為中度加長,但大致比其他基礎物種的要長。頸椎椎體側面有大型深度朝前的凹陷,稱為側凹(pleurocoels),在約巴龍和巴塔哥尼亞龍亦有發育。與約巴龍不同的是,側凹並未被斜骨脊分隔。中線龍骨突出現於椎體前端下側,鯨龍缺乏此一構造。前關節突尖端有個三角形延伸,亦可見於約巴龍,雖然後者的較深。後關節突之上有個相對較大的上骺,為供肌肉附著的骨質突起。頸椎與南非和印度的基礎蜥腳類有所不同。橫關節突(神經弓的側向突起)略傾斜向下,並在前緣有個三角形輪廓-這是在南方物種中沒有的特徵。此外,神經棘邊緣和前表面有皺褶,靠近頸部基部處的側面視角較寬、高度低。椎體與神經弓之間側視可見U字形的凹陷,是棘刺龍的一項自衍徵。[1]
胸椎不尋常地有個駝狀內部結構(數個小型中空腔室),此特徵也在後來的巨龍形類、馬門溪龍科看到,是為趨同演化。[26]雖然前部胸椎椎體有深側凹,這些凹陷沿著軀幹向後逐漸變淺。最後面的胸椎比例上較短。相較之下,杏齒龍和巴塔哥尼亞龍的最後胸椎家長並有明顯的側凹。胸椎神經通道非常狹窄但高聳。[1]全部胸椎都有下椎弓突-下椎弓凹關節(副關節突),使脊柱更僵硬。[1][27]神經棘前後側有皺褶,如同其他基礎蜥腳類。最前面的尾椎神經棘也有與胸椎相同的皺褶,只有峨嵋龍也有此特徵。尾部後部神經棘大幅向後傾斜延伸到其後續尾椎前部,與一些東亞蜥腳類、巨腳龍、約巴龍相似。[1]
第二至五胸椎的肋骨平坦並朝向後方,第六至十一胸椎橫切面更為圓潤並朝向垂直。於是可將肋腔清楚區分為胸段和腰段;相同特徵只有另一種蜥腳類中有敘述過-叉龍科的短頸潘龍。此外胸肋末端還有連接腹肋的附著處。尾部前部人字骨(椎體下方的對稱骨頭)呈刀片狀,是一項基礎特徵。尾部後部人字骨呈桿狀,左右部分彼此分離;此桿狀人字骨很可能緊密附著於椎體底緣;在椎體長邊中段處與前後接續的人字骨關節相連,支撐著椎關節,限制了尾部的彎曲。[1]
肢帶、肢骨及錯誤鑑定的尾刺
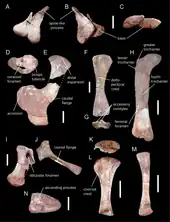
肩胛骨大幅彎曲,下端擴張成扇形,在蜥腳類中相當獨特。其橫切面呈D字形,是真蜥腳類的典型特徵。上端寬闊,下緣有個骨質凸緣。[19]肩胛骨這些特點與亞洲的馬門溪龍科相似,但與岡瓦納物種如火山齒龍、巨腳龍、巴塔哥尼亞龍不同,牠們的下端只有輕微擴張並缺乏後部凸緣。與肩胛骨下端關節連接的鳥喙骨呈現獨特的腎形,被認為是個自衍徵。鳥喙骨有個大的肱二頭肌結節,是肱二頭肌的附著處。胸骨厚實,但較約巴龍纖細。前肢只有肱骨保存下來。其下端不對稱,並具有加大的附後髁(骨頭前下緣朝前的突起)-是一項只在馬門溪龍科發現過的特徵。骨盆的恥骨和坐骨厚實,後者末端寬闊。上股骨的特徵在於上端具有小轉子(前後向牽動後腿肌肉附著的骨突)。棘刺龍的第四轉子(固定肌肉向後牽動後腿的背面突起)特別大。靠近第四轉子處有個大型孔洞,在其他蜥腳類均缺乏,故為自衍徵。脛骨與其他基礎蜥腳類相似,而腓骨厚實。腳踝距骨上側的脛腓骨關節面未被骨壁所分隔,有多達八個培養管(血管進入骨頭的開孔)。[1]
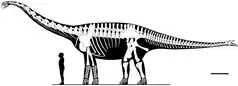

最初被解釋為左右皮內成骨的化石與正模標本一同發現。這些骨頭有個圓狀基部,從中突出一個刺狀突起;內表面為皺褶且凹形。雖然是在骨盆區域發現,研究人員認為在動物生前是長在尾部尖端,而被當作一項鑑定特徵。這項定位是根據左右部分被發現時彼此靠近,代表來自身體中線附近。此外尾部後部因加長的人字骨而變得僵硬,這在其他具有尾槌或尾刺的恐龍也可觀察到。在有親緣關係的蜀龍身上也有相似的尖刺,是構成尾槌的一部分;但棘刺龍可能沒有這樣的尾槌,因為最後幾節尾椎已經太小。可能屬右側的皮內成骨大於左側,形狀略有差異,代表它們沒有成對;可能僅是彼此鏡像反轉對應的情況。而這些差異代表原本是有兩對棘刺。[1]
2013年古生物學家伊曼紐爾·喬普(Emanuel Tschopp)和奧塔維歐·馬締烏斯(Octávio Mateus)重新檢驗了棘刺,發現它們並未具有在其他裝甲恐龍看到的皮內成骨典型的褶皺表面,或是蜀龍擁有的尾槌狀擴張。因為邊緣碎裂,他們也懷疑這些骨頭與過去評估的尺寸有差異。因為這些骨頭是在肩胛骨底下發現,他們表示可能屬於鎖骨,應對這方面的化石進行重新檢驗。[18]Vidal等人於2015年和2018年同意此論點。[28][19]
分類
Remes和同事最初的系統發生學分析認為棘刺龍位於已知最基礎的蜥腳類位置,只比火山齒龍、鯨龍、塔鄒達龍進階,並與真蜥腳類(包含所有較進階的蜥腳類)互成姊妹群。作者表示支持此基礎位置的立場十分薄弱,並討論其他數種位於真蜥腳類中的版本,以解釋非洲和勞亞大陸與其他蜥腳類間的相似性。[1]後續數個研究同樣認為位於真蜥腳類之外的基礎位置,[29][30][31]將棘刺龍列為塔鄒達龍[29]或弗克海姆龍[30]的姊妹分類單元。在一份2013年的論文摘要中,古生物學家裴卓·摩丘(Pedro Mocho)和同事透過整合新清修出來的骨骼所提供的進一步資訊,重新評估了棘刺龍的親緣關係,反駁道棘刺龍坐落於真蜥腳類之內。根據這項分析,棘刺龍比蜀龍、巨腳龍更衍化,並與巴塔哥尼亞龍、馬門溪龍科近緣。[26]2015年的一項研究也提出位於真蜥腳類中的更進階位置,發現棘刺龍與雲龍互成姊妹群。[32]
Nair和Salisbury(2012)的支序圖,支持棘刺龍的基礎位置:[29]
| 沉重龍類 Gravisauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
邢立達和同事(2015)的支序圖,支持位於真蜥腳類中的較進階位置:[32]
| 蜥腳下目 Sauropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
演化

棘刺龍作為已知最完整的基礎蜥腳類之一,為該類群的早期演化及古生物地理學提供了一絲線索。這方面過去所知甚少,因為除了亞洲以外,其他地方的侏羅紀早中期遺骸皆很稀少。Remes等人發現棘刺龍與侏羅紀中期的亞洲蜥腳類有許多共同特徵(尤其是頸椎、尾椎、肩胛骨、肱骨),但與侏儸紀早中期南美洲和印度物種有很大的差異(包括脊椎特徵的形狀及發育、肩胛骨和肱骨形狀)。他們認為這可能是因為侏儸紀中期勞亞大陸蜥腳類與岡瓦那大陸蜥腳類彼此間存在著地理障礙而有隔閡。先前認為在侏儸紀早期與中期之初,蜥腳類平均分布於盤古大陸,多樣性較低,直到盤古大陸在侏羅紀晚期分裂後,才受到地理隔離分化。棘刺龍顯示,過去一度認為在東亞蜥腳類身上觀察到的衍化特徵,其實是真蜥腳類中相對原始的祖徵,即曾居住於勞亞大陸的真蜥腳類保留了在棘刺龍身上也可見到的特徵。而岡瓦那大陸的蜥腳類則構成了一條專行的衍化路線,在地理孤立的過程中已失去了上述祖徵。Remes等人指出,尚需要更多證據來支持這些理論,但可以相信的是,北非、歐洲與東亞的侏儸紀蜥腳類之前的確存在著一定的關聯性。[1]
正如棘刺龍解剖結構與侏儸紀中期蜥腳類分布模式所顯示的,北非可能發生過蜥腳類演化歷程中的重大發展。在侏羅紀早期至中期,當地靠近赤道具有炎熱潮濕的氣候,植物生產力與多樣性皆相當高。另一方面,在侏羅紀早期,南岡瓦那蜥腳類與赤道地區之間由中岡瓦那沙漠區隔開來,使得南北岡瓦那動物群相有所差異。當侏羅紀中期之末沙漠縮減時,新蜥腳類出現並取代了原先典型的侏儸紀中期蜥腳類群。由於北非的約巴龍接近新蜥腳類的基礎位置,且如梁龍超科之類的新蜥腳類在侏羅紀晚期廣泛分布,因此新蜥腳類及其某些分支也可能起源於侏儸紀盤古大陸的赤道地區,當地成為一個蜥腳類演化的「熱點」區域。侏儸紀蜥腳類的多樣化,可能不僅只由地理隔絕所造成,還受到氣候帶和植物地理學的影響。[1]
古生物學
雖然是一群演化成沉重的植食四足動物,棘刺龍和一些蜥腳類卻不如預期地前庭系統(內耳控制平衡和方向的感覺系統)沒有退化。為何棘刺龍會保留此特徵的原因尚未知曉,但假設蜥腳形亞目迷路尺寸和形態可能與頸部長度和活動性有關。前庭系統的擴張可能代表著視覺和雙眼協調、頭頸部活動力的重要性,但對於蜥腳類前庭特徵尚未有確定的解釋。[23]
班傑明·簡根·塞斯基諾(Benjamin Jentgen-Ceschino)等人在2018年的會議中記載了放射狀纖維薄層骨(radial fibrolamellar bone,RFB),是種以放射向管道為特徵的骨骼組織,位於棘刺龍成年標本和伊森龍亞成年標本肱骨皮質(骨頭最外層)的最外層。通常蜥腳類長骨皮質則是纖維薄層複合體。RFB被認為是快速生長的骨骼組織,過去未在蜥腳類發現過。存在於皮質最外層很不尋常,因為通常預期都是長在皮質最內側(對應幼體成長)。棘刺龍的RFB埋於外皮層,並由正常的纖維薄層複合體所接續,代表牠在快速生長後存活了一段時間,這可能反映出受傷而導致的病理性生長。[33]
活動性
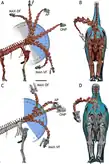
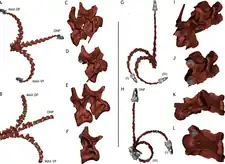
因為棘刺龍是已知最完整的基礎蜥腳類之一,牠成為生物力學研究很好的材料,有助於了解蜥腳類生物學和解剖特徵的功能。Vidal等人(2015)處理3D模型的研究顯示,在受到骨骼形態限制之前,前20節尾椎每個都可偏轉高達20度、側向彎曲8至10度。動物生前還有軟組織和人字骨等限制因素,所以實際活動範圍更小。前20節尾椎亦朝末端變長,如同在梁龍超科所觀察到的情況,並由加大的外軸肌肉提供動力,如大人字骨及第一尾椎角度向上的橫突所示。尾部後部的活動受人字骨重疊所限制,如同馳龍科和甲龍科的情況。綜述以上尾部特徵可能是某種特殊功能的一部分,目前尚未知曉。[28]Joger等人(2009)表示假設的棘刺在尾部末端構成尾刺,可能作為防禦掠食者的武器。[2]
2017年約翰·弗米諾斯(John Fronimos)與傑弗瑞·威爾森(Jeffrey Wilson)使用棘刺龍作為範本來研究蜥腳類神經中樞縫線的複雜性如何對脊椎強度產生影響。蜥腳類同時透過快速與長期生長達到正常的巨大體型。只要個體仍處於成長階段,神經中樞縫線就以軟骨結合的形式來確保成長。一旦達到完全成長體型,軟骨就變成骨頭,將縫線封閉起來。但軟骨可能對神經弓移動(如彎曲和扭轉)壓力承受度較低許多;這在大體型變得格外重要。蜥腳類和其他主龍形類透過提升縫線複雜度來彌補這種結構弱勢,代表神經弓和椎體連結面具有複雜、互相鎖緊的溝脊。在棘刺龍中,縫線複雜度在軀幹前部最明顯,表示該區域壓力最大,可能是長頸部和肋骨重量所造成。朝頭骨和薦骨兩側方向複雜度逐漸降低。稜嵴方向能夠鑑定出脊椎受的壓力類型:頸椎的稜嵴主要防止神經弓前後向移位,而身體脊椎的稜嵴對於防止扭轉更有效率。[34]
2018年的會議摘要中Vidal使用虛擬棘刺龍骨架來檢驗蜥腳類的假想求偶姿勢,為泄殖腔之吻(cloacal kiss)而非使用插入器,如「將腿跨坐背上」、雄性從背後騎上站立或躺下的雌性、「背對背交配」等假設。排除掉會造成骨骼脫位的姿勢:將腿跨坐背上首先排除,因站立的雄性不可能將腿伸過蹲下雌性的背部和臀部,就算後者體型小於前者(目前也沒有證據顯示蜥腳類在體型上有兩性異形)。雄性從雌性背後騎上,並將前腿跨在後者背上的姿勢被認為可行(尾部靈活度足以迴避),但即使雌性躺得再低,彼此泄殖腔也不夠接近到可以相吻。泄殖腔之吻只有透過背對背交配的方式才有可能達成,雄性和雌性彼此向背後靠近,尾部側撇來連接泄殖腔,且四肢的活動性允許體型不同的個體用此方式交配。背對背交配是蜥腳類唯一可能不用透過插入器達成交配的姿勢。這種姿勢在節肢動物、肉食性哺乳類(如犬屬)交配時的「交纏」現象也很普遍。[35]
2020年Vidal等人使用成體和幼體的3D模型來測量頸部活動範圍(靈活度)。此估計假設脊椎間的原始距離能夠可靠的預測,且關節突始終保持接觸。Vidal等人證明以上兩種假設在現代長頸鹿得到佐證,從而增加了對絕種動物運動範圍估計的信心。隨著棘刺龍成長,頸部活動範圍也逐漸增加(更陡峭的頸部姿勢成為可能);脊椎之間的空隙變大;頸部在自然姿勢時變得更加傾斜。長頸鹿生長過程亦觀察到同樣現象。由於棘刺龍脊椎數目較多,就算單個關節彈性遠不及長頸鹿,頸部彈性仍能與後者一樣靈活。棘刺龍可能以與長頸鹿相似的姿勢覓食,並可能是適應高處攝植的蜥腳類中最原始的。高處攝植亦可由解剖構造得到支持,包括狹窄的吻部、寬闊的牙齒、肱骨對肩胛骨比例較長。如同長頸鹿,棘刺龍幼體和成體皆無法僅讓頸部降低接近地面,而可能需要喝水時向兩側伸展前肢。睡覺時長頸鹿將頸部朝身體側向彎曲;雖然棘刺龍頸椎關節本來就夠彎曲,但加長頸肋卻可能限制了彎曲。[15]同年Vidal等人補充說明數位模型揭露的較垂直姿勢及更大範圍的上下運動也支持了棘刺龍的高處攝植能力。很顯然地,高處攝植是真蜥腳類的基礎特徵,而非蜥腳類的蜥腳形亞目的身體比例則顯示牠們是在中度攝植。[22]
古生態學


棘刺龍來自尼日的艾爾哈澤頁岩層,根據紅色黏土顯示出互層砂岩床與地表暴露和恐龍足跡的痕跡,鑑定這裡屬於艾爾哈澤群基部,地層上位於泰加瑪群(Tegama Group)露頭之下。艾爾哈澤層早期被認為年代介於侏儸紀晚期與白堊紀最早期交界,因為上覆的條拉輪組一度被認為年代為下白堊紀。後續研究發現後者其實是侏儸紀中期至晚期之初。棘刺龍的地層學和種系發生學位置(與全世界其他基礎蜥腳類比較)指出可能來自侏儸紀中期(巴柔階至巴通階)。有鑑於艾爾哈澤群尚無法直接進行定年檢測,也有可能早到侏儸紀早期。最底下的交界由下層的阿加德茲群(Agadez Group)所確定,屬於三疊紀晚期(根據手獸足跡化石來判斷)。[1][36]

棘刺龍骨骼發現於粗粒徑至細層壓的紅色粉砂岩,基質中含有一些碳酸鹽。化石採集於數公尺厚的粉砂岩層上半部。當地岩層為近水平向,夾帶小斷層,頂部(約高於骨骼一公尺處)普遍含有古土壤和碳酸鹽。[1]艾爾哈澤群沉積岩是在艾爾勒梅登盆地(與鄰近盆地接壤到古大西洋海灣)還是大河谷系統一部分的時候、河相-湖相作用(與河流湖泊有關聯)的條件下沉積而成的。[36][37]
有四顆獸腳類牙齒被發現與棘刺龍正模標本緊密關聯(在脊椎、恥骨、髖臼處);其中三顆與斑龍科和異特龍科相似,而第四顆屬於認為已知最古老的棘龍科之一。雖然棘刺龍本身骨骼保存良好,牙齒卻保存很差的事實代表著,牙齒是在與蜥腳類一同掩埋前被運輸過來的。[38]在棘刺龍位置以外數百公尺處發現了一些恐龍足跡,包含6個中型蜥腳類足跡步道、以及120個保存良好的二趾獸腳類足跡。二趾足跡起初認為由近鳥類恐龍(如以二趾步行的馳龍科、傷齒龍科)所留下,但後來被解釋為獸腳類游泳時所產生的(解釋了為何有一趾沒留下痕跡)。[36][39]
參考來源
- Remes, K.; Ortega, F.; Fierro, I.; Joger, U.; Kosma, R.; Marín Ferrer, J. M.; Ide, O. A.u; Maga, A.; Farke, A. A. . PLOS ONE. 2009, 4 (9): e6924. Bibcode:2009PLoSO...4.6924R. PMC 2737122
 . PMID 19756139. doi:10.1371/journal.pone.0006924.
. PMID 19756139. doi:10.1371/journal.pone.0006924. - Joger, U.; Kosma, R.; Krüger, F. J. . Braunschweig: Cargo Verlag. 2009. ISBN 978-3-938693-17-9 (德语).
- Ortega, F.; Fierro, I.; Chiappe, L.; et al. . First International Congress on North African Vertebrate Palaeontology (NAVEP1). 25–27 May 2009, Marrakech – Morocco: 49–51. 2009.
- Joger, U. . Google Arts & Culture. [20 April 2019]. (原始内容存档于20 April 2019).
- Joger, U. . Google Arts & Culture. [20 April 2019]. (原始内容存档于20 April 2019).
- Joger, U. . Google Arts & Culture. [20 April 2019]. (原始内容存档于20 April 2019).
- Joger, U. . Beck, L. A. (编). . Natural History Collections. Springer Publishing. 2018: 183–196. ISBN 978-3-319-44321-8. doi:10.1007/978-3-319-44321-8_15.
- Kosma, R. . Google Arts & Culture. [20 April 2019]. (原始内容存档于20 April 2019).
- Witzmann, F.; Hampe, O.; Rothschild, B. M.; Joger, U.; Kosma, R.; Schwarz, D.; Asbach, P. . Journal of Vertebrate Paleontology. 2016, 36 (2): e1080719. doi:10.1080/02724634.2016.1080719.
- Kosma, R. . Beck, L. A.; Joger, U. (编). . Natural History Collections. Springer. 2018: 81–92. ISBN 978-3-319-77401-5. doi:10.1007/978-3-319-77401-5_9.
- Ritter, A. . Google Arts & Culture. [20 April 2019]. (原始内容存档于20 April 2019).
- Vidal, D.; Aberasturi, A.; Mocho, P.; Ortega, F.; Sanz, J. L. . . Burgos: 147–148. 2016.
- . AQUÍ en Elche. 2018 [15 June 2019] (西班牙语).
- García-Martínez, D.; Vidal, D.; Ortega, F. . 16th European Association of Vertebrate Paleontologists Meeting, Caparica (Portugal). European Association of Vertebrate Palaeontologists: 75. 2018.
- Vidal, Daniel; Mocho, Pedro; Páramo, Adrián; Sanz, José Luis; Ortega, Francisco. . PLOS ONE. 2020-01-13, 15 (1): –0227537 [2020-01-13]. Bibcode:2020PLoSO..1527537V. ISSN 1932-6203. PMC 6957182 . PMID 31929581. doi:10.1371/journal.pone.0227537. (原始内容存档于2021-08-14).
- Timmons, J. . Mostly Mammoths, Mummies and Museums. 2018 [24 February 2019]. (原始内容存档于2 March 2019).
- Holtz, T. R. (PDF). 2011 [13 January 2012]. (原始内容存档 (PDF)于2017-08-12).
- Tschopp, E.; Mateus, O. . Journal of Anatomy. 2013, 222 (3): 321–340. PMC 3582252 . PMID 23190365. doi:10.1111/joa.12012.
- Mocho, P.; Vidal, D.; Aberasturi, A.; Kosma, R.; Ortega, F. . 16th European Association of Vertebrate Paleontologists Meeting, Caparica (Portugal). European Association of Vertebrate Palaeontologists: 124. 2018.
- Blázquez, A. P.; Ortega, F. . 10th European Association of Vertebrate Paleontologists Meeting, At Teruel (España). 2012.
- Paul, G. S. . Princeton University Press. 2016: 196. ISBN 978-0-691-16766-4.
- Vidal, D.; Mocho, P.; Aberasturi, A.; Sanz, J. L.; Ortega, F. . Scientific Reports. 2020, 10 (1): 6638. Bibcode:2020NatSR..10.6638V. PMID 32313018. doi:10.1038/s41598-020-63439-0.
- Knoll, F.; Witmer, L. M.; Ortega, F.; Ridgely, R. C.; Schwarz-Wings, D. Farke, A. A. , 编. . PLOS ONE. 2012, 7 (1): e30060. Bibcode:2012PLoSO...730060K. PMC 3260197 . PMID 22272273. doi:10.1371/journal.pone.0030060.
- Taylor, M. P. . PeerJ PrePrints. 2015. doi:10.7287/peerj.preprints.1418v1.
- Brett-Surman, M.K.; Holtz, Jr., Thomas R.; Farlow, James O. . . Indiana University Press. 2012: 135–149. ISBN 978-0-253-00849-7.
- Mocho, P.; Ortega, F.; Aberasturi, A.; Escaso, F. . . Salas de los Infantes Burgos: 89–90. 2013.
- Apesteguía, S. . Tidwell, V.; Carpenter, K. (编). . Bloomington and Indianapolis: Indiana University Press. 2005: 248–267. ISBN 978-0-253-34542-4.
- Vidal, D. C.; Ortega, F. . 63rd SVPCA (Symposium for Vertebrate Palaeontology and Comparative Anatomy), At Southampton, England. 2015.
- Nair, J. P.; Salisbury, S. W. . Journal of Vertebrate Paleontology. 2012, 32 (2): 369. doi:10.1080/02724634.2012.622324.
- Holwerda, F. M.; Pol, D. . Spanish Journal of Palaeontology. 2018, 33 (2): 289. doi:10.7203/sjp.33.2.13604.
- Bronzati, M.; Benson, R. B. J.; Rauhut, O. W. M.; Mannion, P. . Palaeontology. 2018, 61 (2): 289–302 [2021-04-23]. doi:10.1111/pala.12344. (原始内容存档于2021-06-23).
- Xing, L.; Miyashita, T.; Currie, P. J.; You, H.; Zhang, J.; Dong, Z. . Acta Palaeontologica Polonica. 2015, 60 (1): 145–155. doi:10.4202/app.2012.0151.
- Jentgen-Ceschino, C.; Stein, K. . 5th International Palaeontological Congress – Pierre and Marie Curie Campus of Sorbonne University; National Museum of Natural History, Paris, France. 2018. doi:10.13140/RG.2.2.10914.27840.
- Fronimos, J. A.; Wilson, J. A. . Ameghiniana. 2017, 54 (1): 36–49. doi:10.5710/AMGH.05.09.2016.3009.
- Vidal, D. C. . XVI EJIP, At Zarautz. 2018.
- Mudroch, A.; Richter, U.; Joger, U.; Kosma, R.; Idé, O.; Maga, A.; Farke, A. A. . PLOS ONE. 2011, 6 (2): e14642. Bibcode:2011PLoSO...614642M. PMC 3038851 . PMID 21339816. doi:10.1371/journal.pone.0014642.
- Moody, R. T. J. . Selley, R. C. (编). 3. Elsevier Science. 1997: 89–103. ISBN 978-0444825711. doi:10.1016/S1874-5997(97)80008-1.
- Serrano-Martínez, A.; Vidal, D.; Sciscio, L.; Ortega, F.; Knoll, F. . Acta Palaeontologica Polonica. 2015: 403–415. doi:10.4202/app.00101.2014.
- Lockley, M. G.; Harris, J. D.; Li, R.; Xing, L. D.; van der Lubbe, T. . P. L. Falkingham; Da. Marty; A. Richter (编). . Bloomington and Indianapolis: Indiana University Press. 2016: 183–200. ISBN 978-0-253-02102-1.