葡萄園龍屬
葡萄園龍(屬名:,發音:/ˌæmpɪloʊˈsɔːrəs/ AM-pi-loh-SOR-əs,意為「藤蔓蜥蜴」)是一屬泰坦巨龍類的蜥腳類恐龍,生存於白堊紀晚期的歐洲,化石發現於法國南部奥德河畔康帕涅附近的下紅泥灰層。模式種奧德河葡萄園龍(Ampelosaurus atacis)於1995年由讓·呂勒福(Jean Le Loeuff)命名、發表。另一個可能的未命名種,將葡萄園龍的生存年代延伸到白堊紀最末期的馬斯垂克階,約7000萬至6600萬年前。
| 葡萄园龙属 化石时期:白堊紀晚期 | |
|---|---|
 | |
| 葡萄園龍骨架模型 展示於法國埃斯佩拉扎恐龍博物館 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | †蜥脚形亚目 Sauropodomorpha |
| 下目: | †蜥腳下目 Sauropoda |
| 演化支: | †泰坦巨龍類 Titanosauria |
| 演化支: | †岩盔龙类 Lithostrotia |
| 科: | †细长龙亚科 Lirainosaurinae |
| 属: | †葡萄园龙属 Ampelosaurus Le Loeuff, 1995 |
| 模式種 | |
| †奧德河葡萄園龍 Ampelosaurus atacis Le Loeuff, 1995 | |
如同多數蜥腳類,葡萄園龍有著典型的長頸子及長尾巴,背部長著由皮內成骨構成的鱗甲,身長約15公尺及體重約15公噸。有超過500塊骨頭被歸入葡萄園龍,其中包括腦殼,因此能有較深入的解剖學研究。牠是白堊紀晚期北半球少數幾種發現化石較多的泰坦巨龍類,且多虧近年來受到的關注,使葡萄園龍成為法國最著名的恐龍之一。
發現

葡萄園龍最初是在法國南部奧德省奧德河畔康帕涅市鎮附近發現的。出土於下紅泥灰層下部層位,地質年代屬於晚白堊世馬斯垂克階早期,約7000萬年前。這些沉積物顯示過去這裡曾是個佈滿河道的氾濫平原。[1]1995年,法國古生物學家讓·呂勒福(Jean Le Loeuff)首次敘述、命名了模式種奧德河葡萄園龍(Ampelosaurus atacis)。屬名源自古希臘語的αμπελος「藤蔓」,象徵著「葡萄園的蜥蜴」,取自發現地南邊附近、里莫氣泡酒(Blanquette de Limoux)產地的葡萄園。種名是奧德河的拉丁化atax,意為「來自奧德河的」。[1]

首批遺骸於1989年出土自一個屍骨層,產出包含大量肋骨、背椎、尾椎以及許多肢骨,但除了牙齒之外沒有發現其他頭骨材料,還發現四個不同尺寸和形狀的皮內成骨。這些化石分屬於許多隻不同個體。自1989年以來,在法國同一地區相繼發現了更多化石,包含相對完整的骨架與一些頭骨及下頜材料。[2]
後來在2001年,當初的奧德河畔康帕涅化石點由一匹新人所再次找到。他們經過長達13年的搜索,終於發現了一具幾乎完整的骨架,其中涵蓋更多原本正模標本所缺乏的骨骼部位。化石後來全部運往埃斯佩拉扎的恐龍博物館保存,前後耗時近10年來清修與裝設骨架。[3]
另一個未命名物種於2007年在西班牙馬德里-瓦倫西亞高速鐵路建設工程期間,在卡斯提爾-拉曼查村莊附近,屬於原本就盛產許多動物化石的維亞巴德拉席耶拉層,一個稱為羅韋科(Lo Hueco)地點發現。在幾個月有限的時間內,60多名古生物學家和100名工人合作展開了大規模緊急挖掘活動,救回數千個坎潘階晚期至馬斯垂克階早期的植物、無脊椎動物、脊椎動物化石標本,其中包含葡萄園龍未命名種。[4]
敘述
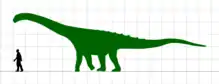
如同許多蜥腳類,葡萄園龍具有典型的長頸部和長尾部,背部還具有由25至28公分長的皮內成骨所構成的裝甲。發現了四個皮內成骨分別展現了三種不同的形態:板狀、球狀、棘刺狀。葡萄園龍估計身長約15至16公尺,體重約15000公斤。[1][5]對於奧德河葡萄園龍的了解來自大量零散、關連的骨骼及牙齒。牠是來自法國本土知名度最高的恐龍之一。[1]科里亞等人(2015)曾提出葡萄園龍可能是某種島嶼侏儒化的結果,因為體型明顯小於其他泰坦巨龍類祖先。[6]
骨骼

葡萄園龍是歐洲已知最完整的泰坦巨龍類蜥腳類之一。自1989年起,在法國南部貝里尤(Bellevue)地點發現超過500具骨骼,包含大量保存良好的材料,已被歸入奧德河葡萄園龍。在初始敘述論文中,只檢驗了一顆牙齒和一些脊椎。2005年呂勒福發表了一篇關於所有已知材料的完整敘述,使得葡萄園龍更深入的被了解。所有材料都被歸入模式種,就算不同的肢骨之間比例上有所差異,相信都只跟個體發育差異有關。[2]

對奧德河葡萄園龍頭部的了解來自少量、但保存良好的牙齒,以及一些頭骨碎片,而在初始論文中描述了其中一顆牙齒。葡萄園龍的牙齒和另外兩種歐洲泰坦巨龍類馬扎爾龍、細長龍有所不同,前者頂部大致呈圓柱形,底部有薄的擴張。牙齒高21毫米、寬6毫米。這些擴張使齒根和齒冠間呈現略微收縮。牙齒也不同於泰坦巨龍類普遍的釘狀齒。[2]牙齒形狀與南極龍還有東哲宮的腦殼有著相似的整體形態。另一個來自南法的頭骨也被描述,但顯示出某些差異,代表白堊紀晚期的法國可能曾存在過至少兩種不同的泰坦巨龍類。[2]
肩胛骨被發現與鳥喙骨連接在一起。骨頭長邊有72公分。肩胛骨片呈三角形,靠近基部處較大,與其他泰坦巨龍類不同。骨片遠端變窄而非擴張。與馬扎爾龍和薩爾塔龍不同的是,肩胛骨在骨片基部缺乏背脊。鳥喙骨呈四邊形,[1]並有增厚的頭緣。[2]
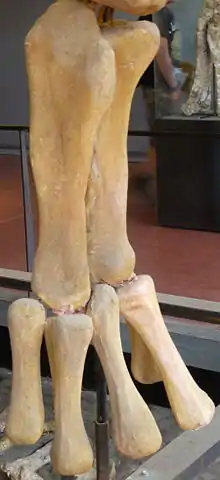
葡萄園龍的骨盆區域也已知。恥骨長約75公分,遠端大幅擴張並有個大孔,類似巴西的泰坦巨龍類。與南美洲泰坦巨龍類相反的是,髂骨側面缺乏擴張。已知的坐骨是個軸,遠端未擴張、發育良好、且非常平坦。[2]已知有個不完整的橈骨,缺乏近、遠末端,有道非常明顯的嵴沿著發育。[2]最大且保存最佳的標本為成對的尺骨。右尺骨全長39.5公分,小而纖細,肘突消失,並有道明顯的嵴。左尺骨較大,長72.5公分。[2]尺骨有個給橈骨的深窩。[7]

貝爾維(Bellevue)約發現了27個股骨,成為最常見的骨骼部位,其中許多或多或少是完整的。頭尾向非常平坦,但另一方面卻缺乏任何不尋常的特徵。[2]一份研究葡萄園龍骨頭生理學的研究發現已知最大股骨長度是110公分,比之前所知的80.2公分還要大許多。這意味著葡萄園龍的祖先型體型比本身還小。[8]已知有18個肱骨,[2]但沒有任何一個尺寸接近股骨那麼大。肱骨結實,長約63公分,近遠末端都大幅擴張,[2]缺乏中空的髓腔。[8]

在貝爾維也發現了三種不同類型的皮內成骨,包括兩種具有大型棘刺的。每個棘刺高約12公分,基部有兩個大孔,每個內面都呈凹凸狀,棘刺反側有一道嵴。側視呈現出兩個區域:逐漸變厚的下半部、棘刺半部。[2]

奧德河葡萄園龍的正模標本是一系列三個背椎構成的脊柱。頸椎在貝爾維很罕見,大多保存狀態都很差。[2]脊椎呈雙後凹(opisthocoelous),椎體向後加長。脊椎神經棘有保存下來,面朝向後方。兩個背椎保存了完整、但嚴重擠壓變形的神經弓、以及典型形狀的神經棘:遠端非常寬,朝下變窄。脊椎內部結構為海綿狀,具有非常大的腔室。[1]尾椎全部強烈雙前凸(procoelous),側邊收縮。尾椎神經棘非常狹窄且高,指向後方。中部尾椎側面比其他還要更為收縮,椎體比例上也較長。[1]貝爾維已知有兩個薦骨,保存很差,被認為屬於幼體。2002年發現了一個完整的薦骨。[2]
鑑定特徵
呂勒福在他的葡萄園龍敘述論文中,列出了以下鑑定特徵來將葡萄園龍與其他蜥腳類區別開來:牙齒的匙狀程度弱;背椎神經棘末端橫向擴張;背椎神經弓強烈向後傾斜;肩胛骨缺乏遠端擴張;肩胛骨頂部有個淺的腹嵴;皮內成骨有板狀、球狀、棘狀形式。[1]而在2005年呂勒福又追加了另外兩項可能的鑑定特徵:背椎神經棘收縮、頸椎也是。[2]
葡萄園龍未命名種

一個來自羅韋科(Lo Hueco)的腦殼化石,編號為MCCM-HUE-8741在一份2013年發表於《PLoS ONE》期刊的論文中假設性的編列為葡萄園龍的一個未命名種(A. sp.)。腦殼被發現與奧德河葡萄園龍有很多共有特徵,例如頭骨後部平坦。腦殼整體尺寸小型,前後長度為10.08公分,左半邊最大寬度為6.43公分。腦殼底半部遺失,儘管如此,標本並未呈現出變形跡象,左右兩邊的形狀沒有太大的差異。[4]
兩個額骨保存下來。長5.73公分及寬6.43公分。每個額骨上面都不平滑,各有一道嵴沿著額骨發育,兩道嵴一同構成眼眶頂。左右頂骨也已發現。彼此連接處由一道ω字形的嵴標示出來。側視頂骨有兩個延生物,這些沒有完整保存下來,但其中部位於上顳孔邊緣上。每個保存的頂骨寬7.96公分。來自羅韋科的基枕骨標本獨特之處在於其枕髁寬度大於高度。枕髁可能因原本覆蓋的軟骨遺失而具有不規則的表面。完整的腦殼在頭骨位置特別低,朝向側邊。枕髁寬2.86公分、高1.58公分。[4]腦殼基部由副基蝶骨(parabasisphenoid)構成。前耳骨(prootic)是一塊高但不長的骨頭,前後長約1.06公分;基蝶骨(basisphenoid)與後蝶骨(laterosphenoid)、頂骨、枕骨主要位在其側。[4]
神經解剖學

與長頸巨龍比較,葡萄園龍未命名種的內耳呈現出更原始的形態。這個特徵可能與頭部旋轉所涉及動作的範圍限制有關。[4]
如同耆那龍和大多數其他非鳥類主龍類,葡萄園龍未命名種的後腦和中腦在內模呈現得相對較不明顯。與TMM 40435標本和少數其他物種如牛津鯨龍參考標本、長頸巨龍相比,小腦觀察不到特徵性的結節構造。如同TMM 40435標本和其他許多主龍類,葡萄園龍未命名種的腦部後面特別狹窄。[4]
大腦皮質由一個顱內腔造成的明顯收縮與腦部其餘部分區隔開來。腦殼皮質區最後部頂部有小型擴張,與耆那龍不同;但在梁龍超科的叉龍和梁龍已知有更大的擴張。在MCCM-HUE-8741標本中,頭蓋骨中間小開孔導致內模腫脹, 暗示著松果系統,正符合松果體的預期位置,介於前腦和中腦之間。[4]
半規管收縮、大幅彎曲。MCCM-HUE-8741標本的半規系統亦顯示出原始的形態,即半規管位彼此接觸。[4]
分類
尾椎特徵和皮內成骨的存在證實葡萄園龍屬於岩盔龍類,這是一群先進的泰坦巨龍類,也包含阿拉莫龍、薩爾塔龍等知名物種。[9]許多其他研究人員曾將葡萄園龍列為泰坦巨龍類、[10]薩爾塔龍科[7]或泰坦巨龍科[1][6][11][12]也許目前尚未能確認葡萄園龍的確切分類位置,但牠具有許多典型特徵,足以判斷的確屬於泰坦巨龍類。[1] 最近迪耶茲迪亞茲(2018)的研究將葡萄園龍和其他坎潘階至馬斯垂克階的法國和西班牙泰坦巨龍類一同組成一個演化支,稱為細長龍亞科。其中葡萄園龍與吉普賽龍互成姊妹群;而整個演化支的位置則介於薩爾塔龍科、風神龍族、隆柯龍類之間。[13]
| 岩盔龍類 Lithostrotia |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
古生物學
發育
葡萄園龍的成長模式似乎與較基礎的蜥腳類很類似。在葡萄園龍發育至一定程度後,骨骼細部組織便開始與基礎蜥腳類產生很大差異。當細部組織變獨特後,葡萄園龍的發育速率即開始減緩。雖然梁龍超科和基礎大鼻龍類也於假定的性成熟後表現出骨骼發育增加及成長速率趨緩的情況,但過程較葡萄園龍來得漸進。其他蜥腳類的發育生長始於性成熟時,但這在葡萄園龍明顯延遲了。葡萄園龍性成熟後的成長周期似乎變短,因為發育過程是由緩慢骨骼建構(而非快速生長)所主導。成長痕跡通常在蜥腳類中很罕見,或有的話也只出現於發育晚期。成長痕跡的缺乏被認為代表著蜥腳類的發育是快速且持續的,並與蜥腳類是透過快速成長而長至巨大體型的理論相輔相成。在任何葡萄園龍骨骼標本都沒有發現成長痕跡,其中卻觀察到明顯趨緩的發育速率及高度的骨骼發育程度,以上特徵通常與成長痕跡的出現有關,因此顯得很不尋常。[8]
組織學
外部基礎系統(external fundamental system,EFS)是種能在骨頭表面觀察到的特徵,被認為是一層生長終止線,代表個體已經長到最大體型和骨骼成熟度。EFS在多數非泰坦巨龍類蜥腳類都有發現,而葡萄園龍尚未得知。近緣的馬扎爾龍、細長龍(與薩爾塔龍科近緣)、阿拉莫龍(薩爾塔龍科)、布萬龍(非常基礎的泰坦巨龍類)都被發現缺乏EFS,但基礎大鼻龍類歐羅巴龍有,因此可推測缺乏情況可能侷限於泰坦巨龍類。葡萄園龍的骨頭外緣(即EFS的預期位置)保存良好,仍缺乏任何化石證據顯示EFS是否存在。然而在發現缺乏EFS的《PLoS ONE》研究中,沒有檢驗較大的標本,因此葡萄園龍具有EFS的可能性仍不能被排除。有鑒於梁龍超科和基礎大鼻龍類都有EFS,這項特徵也可能存在於泰坦巨龍類。大部分葡萄園龍標本都被發現骨頭內部區域有次級骨化密集排列、外部區域則有少數。葡萄園龍的骨化方向不只面向整個骨頭,這在新蜥腳類中並非典型特徵。此外葡萄園龍的任何骨頭都沒有髓腔。根據骨頭組織學分析,葡萄園龍似乎在其生命史中經歷許多種發育階段。[8]
參考來源
- Le Loeuff, J. (PDF). Comptes Rendus de l'Académie des Sciences. IIa. 1995, 321: 693–699 [2014-01-18]. (原始内容存档 (PDF)于2013-12-30).
- Le Loeuff, J. . Tidwell, V.; Carpenter, K. (编). . Bloomington: Indiana University Press. 2005: 115–137. ISBN 978-0-253-34542-4.
- Souillat, C. & Le Loeuff, J. . La Lettre de Dinosauria. 2002: 1–23.
- Knoll, F.; Ridgely, R. C.; Ortega, F.; Sanz, J. L.; Witmer, L. M. Butler, Richard J , 编. . PLOS ONE. 2013, 8 (1): e54991. Bibcode:2013PLoSO...854991K. PMC 3552955
 . PMID 23355905. doi:10.1371/journal.pone.0054991 .
. PMID 23355905. doi:10.1371/journal.pone.0054991 . - Paul, G.S., 2010 The Princeton Field Guide to Dinosaurs, Princeton University Press p. 207
- Salgado, L.; Coria, R.A. . Carpenter, K.; Tidwell, V. (编). . Bloomington: Indiana University Press. 2005: 430–453. ISBN 978-0-253-34542-4.
- Wilson, J.A. (PDF). Zoological Journal of the Linnean Society. 2002, 136 (2): 217–276. doi:10.1046/j.1096-3642.2002.00029.x .
- Klein, N.; Sander, P. M.; Stein, K.; Le Loeuff, J.; Carballido, J. L.; Buffetaut, E. Farke, Andrew A , 编. . PLOS ONE. 2012, 7 (5): e36907. Bibcode:2012PLoSO...736907K. PMC 3353997 . PMID 22615842. doi:10.1371/journal.pone.0036907 .
- Weishampel, D.B.; Dodson, P.; Osmolska, H. 2nd. Berkeley: University of California Press. 2004: 259–322, 588–593. ISBN 978-0-520-24209-8.
- Wilson, J.A.; Sereno, P.C. . Journal of Vertebrate Paleontology. 1998, 18: 1–68. doi:10.1080/02724634.1998.10011115.
- Le Loeuff, J. [The dinosaurs of the Upper Cretaceous of the Ibero-Armorican Island]. de Carvalho, A.M.G.; Andrade, A.M.; dos Santos, V.F.; Cascalho, J.; Taborda, R. (编). . Lisboa: Museu Nacional de História Natural. 1998: 49–64.
- Le Loeuff, J.; Buffetaut, E. . The Dinosaur Society UK Quarterly. 1996, 1 (4): 1–2.
- Díez Díaz, V.; Garcia, G.; Pereda-Suberbiola, X.; Jentgen-Ceschino, B.; Stein, K.; Godefroit, P.; Valentin, X. . Cretaceous Research. 2018, 91: 429–456. doi:10.1016/j.cretres.2018.06.015.