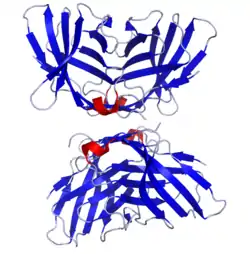

Cytotoxic T-lymphocyte associated protein 4, (CTLA-4) also known as CD152 (cluster of differentiation 152), is a protein receptor that functions as an immune checkpoint and downregulates immune responses. CTLA-4 is constitutively expressed in regulatory T cells but only upregulated in conventional T cells after activation – a phenomenon which is particularly notable in cancers.[5] It acts as an "off" switch when bound to CD80 or CD86 on the surface of antigen-presenting cells. It is encoded by the gene CTLA4 in humans.
The CTLA-4 protein is encoded by the Ctla-4 gene in mice.[6][7]
History
CTLA-4 was first identified in 1991 as a second receptor for the T cell costimulation ligand B7.[8] In November 1995, the labs of Tak Wah Mak and Arlene H. Sharpe independently published their findings on the discovery of the function of CTLA-4 as a negative regulator of T-cell activation, by knocking out the gene in mice.[9][10] Previous studies from several labs had used methods which could not definitively define the function of CTLA-4, and were contradictory.[11]
Function
CTLA-4 is a member of the immunoglobulin superfamily that is expressed by activated T cells and transmits an inhibitory signal to T cells. CTLA-4 is homologous to the T-cell co-stimulatory protein, CD28, and both molecules bind to CD80 and CD86, also called B7-1 and B7-2 respectively, on antigen-presenting cells. CTLA-4 binds CD80 and CD86 with greater affinity and avidity than CD28 thus enabling it to outcompete CD28 for its ligands. CTLA-4 transmits an inhibitory signal to T cells,[12][13][14][9] whereas CD28 transmits a stimulatory signal.[15][16] CTLA-4 is also found in regulatory T cells (Tregs) and contributes to their inhibitory function. T cell activation through the T cell receptor and CD28 leads to increased expression of CTLA-4.
The mechanism by which CTLA-4 acts in T cells remains somewhat controversial. Biochemical evidence suggested that CTLA-4 recruits a phosphatase to the T cell receptor (TCR), thus attenuating the signal.[17] This work remains unconfirmed in the literature since its first publication. More recent work has suggested that CTLA-4 may function in vivo by capturing and removing CD80 and CD86 from the membranes of antigen-presenting cells, thus making these unavailable for triggering of CD28.[18]
In addition to that, it has been found that dendritic cell (DC) - Treg interaction causes sequestration of Fascin-1, an actin-bundling protein essential for immunological synapse formation and skews Fascin-1–dependent actin polarization in antigen presenting DCs toward the Treg cell adhesion zone. Although it is reversible upon T regulatory cell disengagement, this sequestration of essential cytoskeletal components causes a lethargic state of DCs, leading to reduced T cell priming. This suggests Treg-mediated immune suppression is a multi-step process. In addition to CTLA-4 CD80/CD86 interaction, fascin-dependent polarization of the cytoskeleton towards DC-Treg immune synapse may play a pivotal role.[19]
CTLA-4 may also function via modulation of cell motility and/or signaling through PI3 kinase[20] Early multiphoton microscopy studies observing T-cell motility in intact lymph nodes appeared to give evidence for the so-called ‘reverse-stop signaling model’.[21] In this model CTLA-4 reverses the TCR-induced ‘stop signal’ needed for firm contact between T cells and antigen-presenting cells (APCs).[22] However, those studies compared CTLA-4 positive cells, which are predominantly regulatory cells and are at least partially activated, with CTLA-4 negative naive T cells. The disparity of these cells in multiple regards may explain some of these results. Other groups who have analyzed the effect of antibodies to CTLA-4 in vivo have concluded little or no effect upon motility in the context of anergic T-cells.[23] Antibodies to CTLA-4 may exert additional effects when used in vivo, by binding and thereby depleting regulatory T cells.[24]
Structure
The protein contains an extracellular V domain, a transmembrane domain, and a cytoplasmic tail. Alternate splice variants, encoding different isoforms, have been characterized. The membrane-bound isoform functions as a homodimer interconnected by a disulfide bond, while the soluble isoform functions as a monomer. The intracellular domain is similar to that of CD28, in that it has no intrinsic catalytic activity and contains one YVKM motif able to bind PI3K, PP2A and SHP-2 and one proline-rich motif able to bind SH3 containing proteins. The first role of CTLA-4 in inhibiting T cell responses seem to be directly via SHP-2 and PP2A dephosphorylation of TCR-proximal signalling proteins such as CD3 and LAT. CTLA-4 can also affect signalling indirectly via competing with CD28 for CD80/86 binding. CTLA-4 can also bind PI3K, although the importance and results of this interaction are uncertain.
Clinical significance
Variants in this gene have been associated with Type 1 diabetes, Graves' disease, Hashimoto's thyroiditis, celiac disease, systemic lupus erythematosus, thyroid-associated orbitopathy, primary biliary cirrhosis and other autoimmune diseases.
Polymorphisms of the CTLA-4 gene are associated with autoimmune diseases such as rheumatoid arthritis,[25] autoimmune thyroid disease and multiple sclerosis, though this association is often weak. In systemic lupus erythematosus (SLE), the splice variant sCTLA-4 is found to be aberrantly produced and found in the serum of patients with active SLE.
Germline haploinsufficiency
Germline haploinsufficiency of CTLA-4 leads to CTLA-4 deficiency or CHAI disease (CTLA4 haploinsufficiency with autoimmune infiltration), a rare genetic disorder of the immune system. This may cause a dysregulation of the immune system and may result in lymphoproliferation, autoimmunity, hypogammaglobulinemia, recurrent infections, and may slightly increase one’s risk of lymphoma. CTLA-4 mutations have first been described by a collaboration between the groups of Dr. Gulbu Uzel, Dr. Steven Holland, and Dr. Michael Lenardo from the National Institute of Allergy and Infectious Disease, Dr. Thomas Fleisher from the NIH Clinical Center at the National Institutes of Health, and their collaborators in 2014.[26] In the same year a collaboration between the groups of Dr. Bodo Grimbacher, Dr. Shimon Sakaguchi, Dr. Lucy Walker and Dr. David Sansom and their collaborators described a similar phenotype.[27]
CTLA-4 mutations are inherited in an autosomal dominant manner. This means a person only needs one abnormal gene from one parent. The one normal copy is not enough to compensate for the one abnormal copy. Dominant inheritance means most families with CTLA-4 mutations have affected relatives in each generation on the side of the family with the mutation.
Clinical and laboratory manifestations
Symptomatic patients with CTLA-4 mutations are characterized by an immune dysregulation syndrome including extensive T cell infiltration in a number of organs, including the gut, lungs, bone marrow, central nervous system.[28][29] and kidneys. Most patients have diarrhea or enteropathy. Lymphadenopathy and hepatosplenomegaly are also common, as is autoimmunity. The organs affected by autoimmunity vary but include thrombocytopenia, hemolytic anemia, thyroiditis, type I diabetes, psoriasis, and arthritis. Respiratory infections are also common. Importantly, the clinical presentations and disease courses are variable with some individuals severely affected, whereas others show little manifestation of disease. This “variable expressivity,” even within the same family, can be striking and may be explained by differences in lifestyle, exposure to pathogens, treatment efficacy, or other genetic modifiers.[26][27][30][31] This condition is described to have incomplete penetrance of disease. Penetrance is said to be incomplete when some individuals fail to express the trait and seem completely asymptomatic, even though they carry the allele. The penetrance is estimated to be about 60%.
The clinical symptoms are caused by abnormalities of the immune system. Most patients develop reduced levels of at least one immunoglobulin isotype, and have low CTLA-4 protein expression in T regulatory cells, hyperactivation of effector T cells, low switched memory B cells, and progressive loss of circulating B cells.[26][27][31]
Treatment
Once a diagnosis is made, the treatment is based on an individual’s clinical condition and may include standard management for autoimmunity and immunoglobulin deficiencies. A study reported in 2016 treated a Korean CHAI disease patient with abatacept, which is a fusion protein of CTLA-4 and an antibody, and was able to control immune activity and improve patient symptoms. Regular administration of abatacept improved the patient’s severe anemia and diarrhea (3L/day) and brought 3-year-long hospitalization to an end.[31]
Agonists to reduce immune activity
The comparatively higher binding affinity of CTLA-4 than that of CD28 has made CTLA-4 a potential therapy for autoimmune diseases. Fusion proteins of CTLA-4 and antibodies (CTLA4-Ig) have been used in clinical trials for rheumatoid arthritis.[32] The fusion protein CTLA4-Ig is commercially available as Orencia (abatacept). A second generation form of CTLA4-Ig known as belatacept was recently approved by the FDA based on favorable results from the randomized Phase III BENEFIT (Belatacept Evaluation of Nephroprotection and Efficacy as First Line Immunosuppression Trial) study. It was approved for renal transplantation in patients that are sensitized to Epstein–Barr virus (EBV).
Antagonists to increase immune activity
Conversely, there is increasing interest in the possible therapeutic benefits of blocking CTLA-4 (using antagonistic antibodies against CTLA such as ipilimumab—FDA approved for melanoma in 2011—as a means of inhibiting immune system tolerance to tumours and thereby providing a potentially useful immunotherapy strategy for patients with cancer).[5] This therapy was the first approved immune checkpoint blockade therapy.[33] Another is tremelimumab.[5]
The 2018 Nobel Prize in Physiology or Medicine was awarded to James P. Allison and Tasuku Honjo "for their discovery of cancer therapy by inhibition of negative immune regulation".[34]
Interactions
CTLA-4 has been shown to interact with:
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000163599 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000026011 - Ensembl, May 2017
- ↑ "Human PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- ↑ "Mouse PubMed Reference:". National Center for Biotechnology Information, U.S. National Library of Medicine.
- 1 2 3 Syn NL, Teng MW, Mok TS, Soo RA (December 2017). "De-novo and acquired resistance to immune checkpoint targeting". The Lancet. Oncology. 18 (12): e731–e741. doi:10.1016/s1470-2045(17)30607-1. PMID 29208439.
- ↑ Brunet JF, Denizot F, Luciani MF, Roux-Dosseto M, Suzan M, Mattei MG, Golstein P (1987). "A new member of the immunoglobulin superfamily--CTLA-4". Nature. 328 (6127): 267–70. Bibcode:1987Natur.328..267B. doi:10.1038/328267a0. PMID 3496540. S2CID 4316396.
- ↑ Dariavach P, Mattéi MG, Golstein P, Lefranc MP (December 1988). "Human Ig superfamily CTLA-4 gene: chromosomal localization and identity of protein sequence between murine and human CTLA-4 cytoplasmic domains". European Journal of Immunology. 18 (12): 1901–5. doi:10.1002/eji.1830181206. PMID 3220103. S2CID 34071559.
- ↑ Bashyam, Hema (June 2007). "CTLA-4: From conflict to clinic". J Exp Med. 204 (6): 1243. doi:10.1084/jem.2046fta. PMC 2118622. PMID 17632849.
- 1 2 Waterhouse P, Penninger JM, Timms E, Wakeham A, Shahinian A, Lee KP, et al. (November 1995). "Lymphoproliferative disorders with early lethality in mice deficient in Ctla-4". Science. 270 (5238): 985–8. Bibcode:1995Sci...270..985W. doi:10.1126/science.270.5238.985. JSTOR 2888113. PMID 7481803. S2CID 45993765.
- ↑ Tivol EA, Borriello F, Schweitzer AN, Lynch WP, Bluestone JA, Sharpe AH (November 1995). "Loss of CTLA-4 leads to massive lymphoproliferation and fatal multiorgan tissue destruction, revealing a critical negative regulatory role of CTLA-4" (PDF). Immunity. 3 (5): 541–7. doi:10.1016/1074-7613(95)90125-6. PMID 7584144.
- ↑ Pardoll DM (December 2012). "Immunology beats cancer: a blueprint for successful translation". Nature Immunology. 13 (12): 1129–32. doi:10.1038/ni.2392. PMC 4659410. PMID 23160205.
- ↑ Krummel MF, Allison JP (August 1995). "CD28 and CTLA-4 have opposing effects on the response of T cells to stimulation". The Journal of Experimental Medicine. 182 (2): 459–65. doi:10.1084/jem.182.2.459. PMC 2192127. PMID 7543139.
- ↑ Walunas TL, Bakker CY, Bluestone JA (June 1996). "CTLA-4 ligation blocks CD28-dependent T cell activation". The Journal of Experimental Medicine. 183 (6): 2541–50. doi:10.1084/jem.183.6.2541. PMC 2192609. PMID 8676075.
- ↑ Walunas TL, Lenschow DJ, Bakker CY, Linsley PS, Freeman GJ, Green JM, et al. (August 1994). "CTLA-4 can function as a negative regulator of T cell activation". Immunity. 1 (5): 405–13. doi:10.1016/1074-7613(94)90071-x. PMID 7882171.
- ↑ Harding FA, McArthur JG, Gross JA, Raulet DH, Allison JP (April 1992). "CD28-mediated signalling co-stimulates murine T cells and prevents induction of anergy in T-cell clones". Nature. 356 (6370): 607–9. Bibcode:1992Natur.356..607H. doi:10.1038/356607a0. PMID 1313950. S2CID 4333730.
- ↑ Magistrelli G, Jeannin P, Herbault N, Benoit De Coignac A, Gauchat JF, Bonnefoy JY, Delneste Y (November 1999). "A soluble form of CTLA-4 generated by alternative splicing is expressed by nonstimulated human T cells". European Journal of Immunology. 29 (11): 3596–602. doi:10.1002/(SICI)1521-4141(199911)29:11<3596::AID-IMMU3596>3.0.CO;2-Y. PMID 10556814.
- ↑ Lee KM, Chuang E, Griffin M, Khattri R, Hong DK, Zhang W, et al. (December 1998). "Molecular basis of T cell inactivation by CTLA-4". Science. 282 (5397): 2263–6. Bibcode:1998Sci...282.2263L. doi:10.1126/science.282.5397.2263. PMID 9856951.
- ↑ Qureshi OS, Zheng Y, Nakamura K, Attridge K, Manzotti C, Schmidt EM, et al. (April 2011). "Trans-endocytosis of CD80 and CD86: a molecular basis for the cell-extrinsic function of CTLA-4". Science. 332 (6029): 600–3. Bibcode:2011Sci...332..600Q. doi:10.1126/science.1202947. PMC 3198051. PMID 21474713.
- ↑ Chen J, Ganguly A, Mucsi AD, Meng J, Yan J, Detampel P, et al. (February 2017). "Strong adhesion by regulatory T cells induces dendritic cell cytoskeletal polarization and contact-dependent lethargy". The Journal of Experimental Medicine. 214 (2): 327–338. doi:10.1084/jem.20160620. PMC 5294852. PMID 28082358.
- ↑ Knieke K, Lingel H, Chamaon K, Brunner-Weinzierl MC (2012). "Migration of Th1 lymphocytes is regulated by CD152 (CTLA-4)-mediated signaling via PI3 kinase-dependent Akt activation". PLOS ONE. 7 (3): e31391. Bibcode:2012PLoSO...731391K. doi:10.1371/journal.pone.0031391. PMC 3295805. PMID 22412835.
- ↑ Schneider H, Downey J, Smith A, Zinselmeyer BH, Rush C, Brewer JM, et al. (September 2006). "Reversal of the TCR stop signal by CTLA-4". Science. 313 (5795): 1972–5. Bibcode:2006Sci...313.1972S. doi:10.1126/science.1131078. PMID 16931720. S2CID 27123046.
- ↑ Rudd CE, Taylor A, Schneider H (May 2009). "CD28 and CTLA-4 coreceptor expression and signal transduction". Immunological Reviews. 229 (1): 12–26. doi:10.1111/j.1600-065X.2009.00770.x. PMC 4186963. PMID 19426212.
- ↑ Fife BT, Pauken KE, Eagar TN, Obu T, Wu J, Tang Q, et al. (November 2009). "Interactions between PD-1 and PD-L1 promote tolerance by blocking the TCR-induced stop signal". Nature Immunology. 10 (11): 1185–92. doi:10.1038/ni.1790. PMC 2778301. PMID 19783989.
- ↑ Simpson TR, Li F, Montalvo-Ortiz W, Sepulveda MA, Bergerhoff K, Arce F, et al. (August 2013). "Fc-dependent depletion of tumor-infiltrating regulatory T cells co-defines the efficacy of anti-CTLA-4 therapy against melanoma". The Journal of Experimental Medicine. 210 (9): 1695–710. doi:10.1084/jem.20130579. PMC 3754863. PMID 23897981.
- ↑ Westra HJ, Martínez-Bonet M, Onengut-Gumuscu S, Lee A, Luo Y, Teslovich N, et al. (October 2018). "Fine-mapping and functional studies highlight potential causal variants for rheumatoid arthritis and type 1 diabetes". Nature Genetics. 50 (10): 1366–1374. doi:10.1038/s41588-018-0216-7. PMC 6364548. PMID 30224649.
- 1 2 3 Kuehn HS, Ouyang W, Lo B, Deenick EK, Niemela JE, Avery DT, et al. (September 2014). "Immune dysregulation in human subjects with heterozygous germline mutations in CTLA4". Science. 345 (6204): 1623–1627. Bibcode:2014Sci...345.1623K. doi:10.1126/science.1255904. PMC 4371526. PMID 25213377.
- 1 2 3 Schubert D, Bode C, Kenefeck R, Hou TZ, Wing JB, Kennedy A, et al. (December 2014). "Autosomal dominant immune dysregulation syndrome in humans with CTLA4 mutations". Nature Medicine. 20 (12): 1410–1416. doi:10.1038/nm.3746. PMC 4668597. PMID 25329329.
- ↑ Kaninia S, Grammatikos A, Urankar K, Renowden SA, Patel NK, Gompels MM, Rice CM (August 2021). "CNS demyelination associated with immune dysregulation and a novel CTLA-4 variant". Multiple Sclerosis. 27 (9): 1464–1467. doi:10.1177/1352458520963896. PMC 8358566. PMID 34097529.
- ↑ Grammatikos A, Johnston S, Rice CM, Gompels M (August 2021). "A Family with a Novel CTLA4 Haploinsufficiency Mutation and Neurological Symptoms". Journal of Clinical Immunology. 41 (6): 1411–1416. doi:10.1007/s10875-021-01027-1. PMC 8310858. PMID 33956248.
- ↑ Zeissig S, Petersen BS, Tomczak M, Melum E, Huc-Claustre E, Dougan SK, et al. (December 2015). "Early-onset Crohn's disease and autoimmunity associated with a variant in CTLA-4". Gut. 64 (12): 1889–97. doi:10.1136/gutjnl-2014-308541. PMC 4512923. PMID 25367873.
- 1 2 3 Lee S, Moon JS, Lee CR, Kim HE, Baek SM, Hwang S, et al. (January 2016). "Abatacept alleviates severe autoimmune symptoms in a patient carrying a de novo variant in CTLA-4". The Journal of Allergy and Clinical Immunology. 137 (1): 327–330. doi:10.1016/j.jaci.2015.08.036. PMID 26478010.
- ↑ Westhovens R, et al. (2004). "Abatacept (CTLA4Ig) treatment increases the remission rate in rheumatoid arthritis patients refractory to methotrexate treatment". Arthritis Research & Therapy. 6 (Suppl 1): 86. doi:10.1186/ar1128. PMC 2833769.
- ↑ Pardoll DM (March 2012). "The blockade of immune checkpoints in cancer immunotherapy". Nature Reviews. Cancer. 12 (4): 252–64. doi:10.1038/nrc3239. PMC 4856023. PMID 22437870.
- ↑ "The Nobel Prize in Physiology or Medicine 2018". NobelPrize.org. Retrieved 3 July 2023.
- ↑ Follows ER, McPheat JC, Minshull C, Moore NC, Pauptit RA, Rowsell S, et al. (October 2001). "Study of the interaction of the medium chain mu 2 subunit of the clathrin-associated adapter protein complex 2 with cytotoxic T-lymphocyte antigen 4 and CD28". The Biochemical Journal. 359 (Pt 2): 427–34. doi:10.1042/0264-6021:3590427. PMC 1222163. PMID 11583591.
- ↑ Chuang E, Alegre ML, Duckett CS, Noel PJ, Vander Heiden MG, Thompson CB (July 1997). "Interaction of CTLA-4 with the clathrin-associated protein AP50 results in ligand-independent endocytosis that limits cell surface expression". Journal of Immunology. 159 (1): 144–51. doi:10.4049/jimmunol.159.1.144. PMID 9200449. S2CID 25449038.
- ↑ Peach RJ, Bajorath J, Naemura J, Leytze G, Greene J, Aruffo A, Linsley PS (September 1995). "Both extracellular immunoglobin-like domains of CD80 contain residues critical for binding T cell surface receptors CTLA-4 and CD28". The Journal of Biological Chemistry. 270 (36): 21181–7. doi:10.1074/jbc.270.36.21181. PMID 7545666.
- ↑ Stamper CC, Zhang Y, Tobin JF, Erbe DV, Ikemizu S, Davis SJ, et al. (March 2001). "Crystal structure of the B7-1/CTLA-4 complex that inhibits human immune responses". Nature. 410 (6828): 608–11. Bibcode:2001Natur.410..608S. doi:10.1038/35069118. PMID 11279502. S2CID 4329622.
- ↑ Baroja ML, Vijayakrishnan L, Bettelli E, Darlington PJ, Chau TA, Ling V, et al. (May 2002). "Inhibition of CTLA-4 function by the regulatory subunit of serine/threonine phosphatase 2A". Journal of Immunology. 168 (10): 5070–8. doi:10.4049/jimmunol.168.10.5070. PMID 11994459.
Further reading
- Liossis SN, Sfikakis PP, Tsokos GC (August 1998). "Immune cell signaling aberrations in human lupus". Immunologic Research. 18 (1): 27–39. doi:10.1007/BF02786511. PMID 9724847. S2CID 13581332.
- Chang TT, Kuchroo VK, Sharpe AH (2002). "Role of the B7-CD28/CTLA-4 pathway in autoimmune disease". Signal Transduction Pathways in Autoimmunity. Current Directions in Autoimmunity. Vol. 5. pp. 113–30. doi:10.1159/000060550. ISBN 978-3-8055-7308-5. PMID 11826754.
- Alizadeh M, Babron MC, Birebent B, Matsuda F, Quelvennec E, Liblau R, et al. (July 2003). "Genetic interaction of CTLA-4 with HLA-DR15 in multiple sclerosis patients". Annals of Neurology. 54 (1): 119–22. doi:10.1002/ana.10617. PMID 12838528. S2CID 9216025.
- Chistiakov DA, Turakulov RI (August 2003). "CTLA-4 and its role in autoimmune thyroid disease". Journal of Molecular Endocrinology. 31 (1): 21–36. doi:10.1677/jme.0.0310021. PMID 12914522.
- Vaidya B, Pearce S (May 2004). "The emerging role of the CTLA-4 gene in autoimmune endocrinopathies". European Journal of Endocrinology. 150 (5): 619–26. doi:10.1530/eje.0.1500619. PMID 15132716.
- Brand O, Gough S, Heward J (October 2005). "HLA , CTLA-4 and PTPN22 : the shared genetic master-key to autoimmunity?". Expert Reviews in Molecular Medicine. 7 (23): 1–15. doi:10.1017/S1462399405009981. PMID 16229750. S2CID 841442.
- Kavvoura FK, Akamizu T, Awata T, Ban Y, Chistiakov DA, Frydecka I, et al. (August 2007). "Cytotoxic T-lymphocyte associated antigen 4 gene polymorphisms and autoimmune thyroid disease: a meta-analysis". The Journal of Clinical Endocrinology and Metabolism. 92 (8): 3162–70. doi:10.1210/jc.2007-0147. PMID 17504905.
External links
- Human CTLA4 genome location and CTLA4 gene details page in the UCSC Genome Browser.
- Overview of all the structural information available in the PDB for UniProt: P16410 (Cytotoxic T-lymphocyte protein 4) at the PDBe-KB.
PDB gallery | |
|---|---|
|

This article incorporates text from the United States National Library of Medicine, which is in the public domain.