三羧酸循环
三羧酸循環(tricarboxylic acid cycle) 可簡稱為TCA cycle,亦作檸檬酸循環(citric acid cycle),是有氧呼吸的第二階段。該循環以循環中一個重要中間體檸檬酸命名,又因爲檸檬酸是一種三元羧酸,該反應又稱爲三羧酸循環。該循環亦因由英國生物化學家克雷布斯(Krebs)發現而稱爲克雷布斯循環(Krebs cycle),克雷布斯亦因此項貢獻獲1953年諾貝爾生理學或醫學獎。丙酮酸在經過丙酮酸脫氫酶系氧化,生成乙醯輔酶A(acetyl-CoA)後,與四碳二元羧酸草酰乙酸化合,生成檸檬酸,進入檸檬酸循環。隨後,經過一系列反應,兩個碳原子轉化爲二氧化碳(CO2)分子,檸檬酸中蘊藏的化學能轉化至還原的輔酶中。檸檬酸循環的終產物仍然是草酰乙酸,這使得該循環能源源不斷地氧化輸入循環的乙醯輔酶A。

一般情況下,檸檬酸循環產生的還原輔酶會連同糖酵解過程產生的還原輔酶一同,在氧化磷酸化過程中氧化,生成大量的ATP[1]:148。一分子的乙酰輔酶A在被檸檬酸循環代謝後,可產生兩分子的CO2分子、三分子NADH、一分子FADH2,以及一分子GTP[註 1][2]:340。
檸檬酸循環可以代謝糖類、脂質,以及大部分氨基酸,因爲這三類物質都能轉換爲乙酰輔酶A或檸檬酸循環的中間體,從而進入檸檬酸循環之中。另外,檸檬酸循環的許多中間體可供生物體利用。當中間產物不足時,可通過添補反應對中間產物進行補充。生物體最重要的填補反應是在丙酮酸羧化酶催化下,以一分子丙酮酸和一分子二氧化碳分子爲原料,合成一分子草酰乙酸的反應[2]:343。
發現

1930年代,檸檬酸循環中的一些化學反應就已由阿爾伯特·聖捷爾吉闡明。阿爾伯特·聖捷爾吉因他對檸檬酸循環中的一個重要中間體延胡索酸的研究,而獲得1937年諾貝爾生理學或醫學獎[3]。檸檬酸循環本身則是由德國生物化學家漢斯·阿道夫·克雷布斯在1937年於錫菲大學發現的。他因對檸檬酸循環的研究而獲得1953年諾貝爾生理醫學獎[4]。

進化
構成檸檬酸循環的一些化學反應在厭氧細菌中就已出現了。檸檬酸循環本身則可能經過多次進化[5]。從理論上說,檸檬酸循環並不是沒有相關的替代方案。然而,檸檬酸循環的效率卻是最高的。即使存在進化上獨立的能替代檸檬酸循環的過程,它們也應該已通過趨同演化而與檸檬酸循環融合[6][7]。
在有氧呼吸中的位置
糖類在細胞質基質中發生糖酵解後,生成的終產物是丙酮酸。丙酮酸通過線粒體外膜上的通道蛋白,再通過線粒體內膜上的丙酮酸轉位酶進入線粒體基質。之後,丙酮酸經丙酮酸脫氫酶複合體氧化,轉化爲可進入檸檬酸循環的乙酰輔酶A,這一過程稱爲丙酮酸氧化[2]:332。檸檬酸循環發生於線粒體基質中,但也有一部分會於線粒體內膜或嵴膜上發生。原核生物的檸檬酸循環則發生於細胞質基質中。檸檬酸循環產生的還原輔酶會在氧化磷酸化過程中與氧氣反應,產生的能量會部分轉移到ATP中[8]:368。
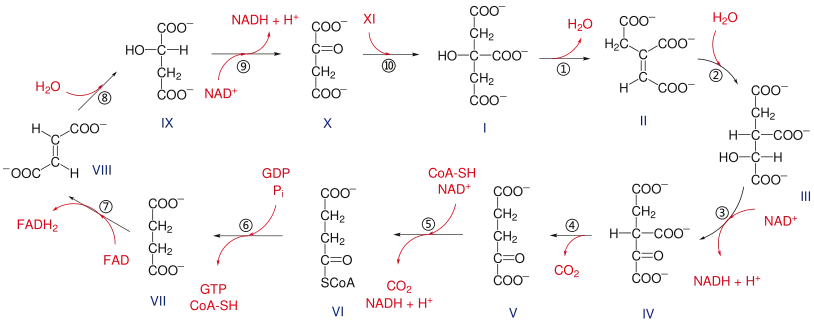
步驟
檸檬酸循環可概括爲8大反應步驟[2]:335[8]:368:
第一步:柠檬酸合酶
.jpg.webp)
柠檬酸合成酶催化乙酰輔酶A(Acetyl-CoA)與四碳的草酰乙酸(Oxaloacetate)发生缩合反应和硫酯水解,生成檸檬酸(Citrate),通常被视为循环的第一步。其四级结构由两个亚基组成,两个底物可以与每个亚基结合。[10]由于硫酯水解释放能量,反应是不可逆的。酶的活性位点激活乙酰辅酶A,使其类似于草酰乙酸的碳中心:沿着两个分子之间的键,硫酯基团(CoA)被水解,形成柠檬酸分子。[11]
该反应高度放能(ΔG°′ = -31.4 kJ/mol),也因此该步骤在标准条件下不可逆。此外,酶产生的柠檬酸能够竞争性抑制酶活性,尽管该反应非常有利(因为是放能的),但柠檬酸合酶可以受到严格调节。[11]例如,柠檬酸盐本身充当柠檬酸合成酶的抑制剂,而草酰乙酸与其结合会增加其对乙酰辅酶A的亲和力。这方面具有相当大的生物学重要性,因为它可以完全调节整个克雷布斯循环,使酶成为整个循环的“起搏器”。[12][13]糖酵解中的磷酸果糖激酶1会被柠檬酸盐抑制,同时它会激活乙酰辅酶 A 羧化酶以进行脂肪酸合成。这一点说明了我们代谢周期的相互关联性。[14]

第二步:烏頭酸酶
.jpg.webp)
檸檬酸在烏頭酸酶(Aconitase)的催化作用下异构化通过生成乌头酸生成異檸檬酸(Isocitrate)。乌头酸酶可以催化順烏頭酸的水合反应及水合产物柠檬酸、异柠檬酸的脱水反应。虽然该反应是可逆的,但在pH=7.0,25攝氏度條件下,平衡混合物中柠檬酸和异柠檬酸的比例为9:1,因爲異檸檬酸不斷消耗,在克雷布斯循环中,由于质量作用定律,反應平衡不斷右移。一旦产生了顺乌头酸,就添加水分子以将双键氧化成羟基,并通过添加水产生异柠檬酸。[16][17]
酶的活性位点有一个铁硫簇,可与一些极性氨基酸残基共同结合底物。丝氨酸、精氨酸、组氨酸和天冬氨酸残基确保与底物的结合,与(1R,2S)-柠檬酸盐单独特异性结合,排斥与其它形式的结合。[18][19][20]

第三步:异柠檬酸脱氢酶
.jpg.webp)
異檸檬酸在異檸檬酸脫氫酶(Isocitrate dehydrogenase)作用下,經中間體草酰琥珀酸(Oxalosuccinate)氧化脱羧生成α-酮戊二酸(α-Ketoglutarate)。线粒体异柠檬酸脱氢酶是一种依赖于NAD+和Mn2+、Mg2+存在的酶,在異檸檬酸轉化爲草酰琥珀酸的過程中,有一分子NAD+還原爲NADH。[22][23]草酰琥珀酸具有β-酮酸的结构,与α位羧基的氧络合的二价离子的存在增加了该区域的电负性,导致分子电子重排,发生脱羧反应(即释放CO2分子),从而形成α-酮戊二酸,末端有两个羧基,和一个酮位于两个羧基之一的α位。该反应中,酶的活性要在Mg2+或Mn2+離子的輔助下才可實現。由于具有足够的放能(ΔG°' = -8.4 kJ/mol),能够将先前的反应从乌头酸酶向前推进。總的來說,該過程生成了一分子NADH以及一分子CO2。[24][25]
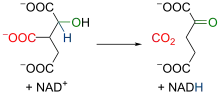
第四步:α-酮戊二酸脱氢酶
异柠檬酸转化为α-酮戊二酸后,α-酮戊二酸在α-酮戊二酸脫氫酶複合體(α-Ketoglutarate dehydrogenase complex)的催化作用下,进行第二次氧化脱羧反应,與輔酶A化合,生成琥珀酰輔酶A(Succinyl-CoA),該過程還會生成一分子CO2,以及一分子NADH。該反應的机理与丙酮酸氧化机理相似,在生理條件下同样是一個單向反應。[27]两个反应都包括α-酮酸的脱羧以及随后与辅酶A生成高能硫酯键,催化这些反应的复合物彼此相似。[28]
α-酮戊二酸脫氫酶複合體由三種酶組成:α-酮戊二酸脫氫酶(E1)、二氫硫辛酰轉琥珀酸酶(E2)、二氫硫辛酰脫氫酶(E3),α-酮戊二酸脫氫酶複合體要在一些輔助因子的輔助下才能發揮功能。该反应的自由能差为ΔG°′ = -30.1 kJ/mol,因此高度放能。[28]

第五步:琥珀酰輔酶A合成酶
.jpg.webp)
琥珀酰辅酶A是一种高能硫酯,其水解ΔG°′ = -33.5 kJ/mol,与ATP的-30.5 kJ/mol相似。柠檬酸合酶利用具有这种高能键的中间体来完成二碳分子(乙酰辅酶A)和四碳分子(草乙酸)之间的融合,而在琥珀酰輔酶A合成酶(Succinyl-CoA synthetase)利用这种能量来磷酸化。
来自硫酯的能量简单地转化为与磷酸键相连的能量:反应的第一步生成高能中间体磷酸琥珀酰,随后,催化位点中存在的组氨酸将磷酸盐从磷酸酯中去除。碳水化合物分子,生成产物琥珀酸和磷酸组氨酸分子,磷酸组氨酸分子迅速将磷酸盐转化为核苷二磷酸盐,将其“充电”为三磷酸盐。
琥珀酸輔酶A轉化爲琥珀酸(Succinate)和輔酶A,生成一分子GTP(該酶有兩個版本,分別生成ATP和GTP;不同生物,或同一生物的不同組織會依對ATP和GTP需求的差異在該步使用相應版本的酶)。这是循环中在底物水平上发生磷酸化、生成高能磷酸鍵的唯一步骤。
接下來,琥珀酸經過三步反應,重新轉化爲草酰乙酸。

第六步:琥珀酸脫氫酶
_(coli).jpg.webp)
循环的最后部分是四碳分子的重新排列,直到草酰乙酸。为实现这一点,琥珀酸上存在的亚甲基桥必须转化为羰基,这种转化通过三个步骤发生:第一次氧化、水合和二次氧化。这三个步骤,除了再生草酰乙酸之外,还可以通过形成FADH2和NADH来提取更多能量。
第一个氧化反应是琥珀酸在琥珀酸脫氫酶(Succinate dehydrogenase)的催化作用下,轉化爲延胡索酸(Fumarate)。該過程中有一分子FAD還原爲FADH2。琥珀酸脱氢酶复合物是循环中唯一以FAD而不是NAD+作为氢受体的酶,FAD通过组氨酸残基与酶共价连接。该酶使用FAD,因为与反应相关的能量不足以还原 NAD+。

第七步:延胡索酸
.jpg.webp)
延胡索酸經延胡索酸酶(Fumarase)催化,催化水分子中的H+和OH-基团加成到反式分子上,由于该酶只能在一侧结合 OH-,轉化爲L-蘋果酸(L-Malate)。
延胡索酶有两类:I类和II类,取决于其相关亚基的排列、对金属的需求及其热稳定性。I类延胡索酸酶在受热或辐射时能够改变状态或变得失活,对超氧阴离子敏感,依赖Fe2+,是二聚体,约120 kD。II类延胡索酸酶存在于原核生物和真核生物中,是 20 kD的四聚酶,包含三个不同的显着同源的氨基酸片段,并且不依赖铁且热稳定。已知原核生物具有三种不同形式的延胡索酸酶:延胡索酸酶A、延胡索酸酶B和延胡索酸酶C,后者属于II类延胡索酶,而延胡索酸酶A和延胡索酸酶B被归类为I类。[32][33]
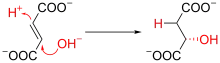
第八步:蘋果酸脫氫酶
.jpg.webp)
L-蘋果酸在蘋果酸脫氫酶(Malate dehydrogenase)的催化作用下,轉化爲草酰乙酸。該過程生成一分子NADH。
与最后一个反应相关的吉布斯自由能显然是正的(与循环中的其他反应不同)。该酶的活性是由柠檬酸合酶消耗草酰乙酸和电子传递链消耗NADH 驱动。該反應平衡傾向於蘋果酸一側,但因草酰乙酸不斷消耗,反應得以持續進行。基於上述理由,該反應也是檸檬酸循環的一個重要調控位點[1]:149。
至此,檸檬酸循環完成。生成的草酰乙酸可再次進入檸檬酸循環[2]:335。

添補反應
三羧酸循環的中間產物可能會因參與其他代謝過程而消耗,添補反應(Anaplerotic reactions)能夠補充檸檬酸循環的各中間產物.使得其物質量保持相對平衡[35]。添補反應中最重要的一個是在丙酮酸羧化酶催化下,以一分子丙酮酸和一分子二氧化碳分子爲原料,合成一分子草酰乙酸的反應。而在植物體內,乙醛酸循環能起到同樣的作用[2]:343。另外,乳酸可以先氧化爲丙酮酸,然後再通過羧化反應轉化爲草酰乙酸進入檸檬酸循環[1]:152。
總反應式
高等動物的檸檬酸循環可用下式表示:[2]:340
乙酰輔酶A+3 NAD+ + FAD + GDP + Pi + 2 H2O →
輔酶A-SH + 3 NADH + 3 H+ + FADH2 + GTP + 2 CO2
調控及抑制
調控
多種途徑均可調控檸檬酸循環的進行。總的來說,當細胞供能充足時(ATP、NADH、乙酰輔酶A等物質濃度升高),檸檬酸循環速度較慢,而當細胞供能處於不足狀態時(ADP濃度高,ATP、NADH濃度低),檸檬酸循環的速度會加快,保證能爲生物體提供適量的ATP以及還原輔酶[8]:370。
首先,因爲檸檬酸循環是呼吸作用的一個環節,對下游的電子傳遞鏈、氧化磷酸化進行調控即可對檸檬酸循環起到調控作用。對檸檬酸循環上游的丙酮酸氧化進行調控也可以起調控檸檬酸活性的作用。另外,也可以通過調控檸檬酸循環中的酶的活性起調控檸檬酸循環的作用。從化學原理上講,對能進行得較徹底的反應進行調控效果較好,因此,丙酮酸脫氫酶、檸檬酸合成酶、異檸檬酸脫氫酶、α-酮戊二酸脫氫酶是較好的調控位點。對檸檬酸循環的調控通常來說與細胞的狀態有密切關係。比如,脫氫酶需要鈣離子的激活,而肌肉收縮時,胞內的鈣離子濃度會升高,這也正是需能較高的時候。此外,一些酶也能直接和ATP/ADP,或NADH/NAD+反應,並發生活性變化,比如異檸檬酸脫氫酶能經由ADP活化,也可透過ATP/NADH來抑制。蘋果酸脫氫酶的活性受NAD+/NADH比值的影響。此外,檸檬酸循環的中間體的濃度也可以對檸檬酸循環的活性起到調控作用。比如,草酰乙酸可以抑制琥珀酰脫氫酶的活性。不過,這類調控在體內到底發揮多大的作用,目前還未得到充分研究。[1]:153[2]:342
抑制
部分物質可以通過化學作用抑制檸檬酸循環的進行,其中,以下三種物質研究得較透徹:氟乙酸、亞砷酸、丙二酸[1]:151。
其中,氟乙酸通過生成氟乙酰輔酶A,再生成氟代乙酰辅酶A进而生成氟檸檬酸。氟檸檬酸能抑制烏頭酸酶的活性,使得檸檬酸循環無法繼續進行。利用這一原理,可以用氟乙酸製造殺蟲劑或滅鼠藥。[2]:337亞砷酸能抑制α-酮戊二酸脫氫酶複合體的酶活性[1]:149。丙二酸與琥珀酸脫氫酶的底物琥珀酸結構相似,能通過與該酶發生不可逆結合使其失活[2]:339。
此外,该途径关键的限速酶柠檬酸合酶受ATP、NADH、琥珀酰CoA的抑制。异柠檬酸脱氢酶受ATP的抑制。α-酮戊二酸脱氢酶系受ATP和琥珀酰CoA的抑制。
與其他代謝過程的關係
檸檬酸循環與包括脂肪酸合成、糖異生、轉氨基作用、嘌呤和嘧啶的合成等在內的多種生物體內的代謝過程密切相關[8]:371。
糖異生及轉氨基作用
在人的肝、腎中,檸檬酸循環中的草酰乙酸可通過糖質新生作用轉化爲葡萄糖。在糖質新生作用中,最關鍵的酶是磷酸丙酮酸羧基激酶,它可以使草酰乙酸脫羧,生成磷酸丙酮酸,GTP分子爲該反應提供能量。值得注意的是,琥珀酰輔酶A轉化爲琥珀酸過程中生成的GTP分子可用於上述過程,是一個糖異生與檸檬酸循環聯繫的紐帶[1]:150。
檸檬酸循環的一些中間體可通過轉氨基作用,經過一系列反應轉化爲氨基酸。比如,α-酮戊二酸可以轉化爲穀氨酸,草酰乙酸可以轉化爲天冬氨酸。上述過程是可逆的,即氨基酸也可以通過脫氨基作用轉化爲檸檬酸循環的中間體而進入檸檬酸循環分解代謝。不過,氨基酸通過上述途徑分解代謝產生的能量很少,一般情況下,只佔生物體總產能的1%不到[2]:438。
注釋
- 部分生物組織在這一步產生的是ATP,而不是GTP。
参考文献
- Robert K. Murray et al. . 科學出版社. 2010. ISBN 978-7-03-028539-3.
- 王鏡岩、朱聖庚、徐長法. . 高等教育出版社. 2008. ISBN 978-7-04-018363-4.
- . The Nobel Foundation. [2011-10-26]. (原始内容存档于2007-02-19).
- . The Nobel Foundation. [2011-10-26]. (原始内容存档于2007-02-22).
- Gest H. . Biochem. Soc. Symp. 1987, 54: 3–16. PMID 3332996.
- Meléndez-Hevia E, Waddell TG, Cascante M. . J. Mol. Evol. September 1996, 43 (3): 293–303. PMID 8703096. doi:10.1007/BF02338838.
- Ebenhöh O, Heinrich R. . Bull. Math. Biol. January 2001, 63 (1): 21–55. PMID 11146883. doi:10.1006/bulm.2000.0197.
- David Hames, Nigel Hooper. . 科學出版社. 2005. ISBN 978-7-03-025218-0 (英语).
- Template:Cita pubblicazione
- Wiegand, Georg; Remington, Stephen J. . Annual Review of Biophysics and Biophysical Chemistry. 1986-06, 15 (1). ISSN 0883-9182. doi:10.1146/annurev.bb.15.060186.000525 (英语).
- Condon, R. . Journal of Gastrointestinal Surgery. 2002-08, 6 (4). ISSN 1091-255X. doi:10.1016/s1091-255x(02)00020-3.
- Wiegand, Georg; Remington, Stephen J. . Annual Review of Biophysics and Biophysical Chemistry. 1986-06, 15 (1). ISSN 0883-9182. doi:10.1146/annurev.bb.15.060186.000525 (英语).
- MacDonald, Michael J; Al-Masri, Hussam; Jumelle-Laclau, Muriel; Cruz, Meredith O. . Diabetes. 1997-12-01, 46 (12). ISSN 0012-1797. doi:10.2337/diab.46.12.1996.
- Bordoni, Bruno; Simonelli, Marta; Morabito, Bruno. . Cureus. 2019-07-23. ISSN 2168-8184. doi:10.7759/cureus.5208.
- Lauble, Hanspeter; Stout, Charles David. . Proteins: Structure, Function, and Bioinformatics. 1995-05, 22 (1). ISSN 0887-3585. doi:10.1002/prot.340220102 (英语).
- Olsen, Luke; Hassan, Huzaifa; Xia, Fanning; Keaton, Sarah; Rohner, Nicolas. . Zoological Research. 2023, 44 (4). ISSN 2095-8137. doi:10.24272/j.issn.2095-8137.2022.486.
- M. DE PANFILIS; L. MARCELLI. . Annals of Geophysics. 2011-05-09, 21 (4). ISSN 2037-416X. doi:10.4401/ag-5072.
- Condon, R. . Journal of Gastrointestinal Surgery. 2002-08, 6 (4). ISSN 1091-255X. doi:10.1016/s1091-255x(02)00020-3.
- Robbins, A. H.; Stout, C. D. . Proteins: Structure, Function, and Bioinformatics. 1989-01, 5 (4). ISSN 0887-3585. doi:10.1002/prot.340050406 (英语).
- Beinert, Helmut; Kennedy, Mary Claire. . The FASEB Journal. 1993-12, 7 (15). ISSN 0892-6638. doi:10.1096/fasebj.7.15.8262329.
- Stoddard, B.L.; Mesecar, A.; Koshland Junior, D.E. . dx.doi.org. 1997-11-12 [2024-01-21].
- Schowen, Richard L. . Journal of Chemical Education. 1993-08, 70 (8). ISSN 0021-9584. doi:10.1021/ed070pa223.1.
- Yasutake, Yoshiaki; Watanabe, Seiya; Yao, Min; Takada, Yasuhiro; Fukunaga, Noriyuki; Tanaka, Isao. . Journal of Biological Chemistry. 2003-09, 278 (38). doi:10.1074/jbc.M304091200 (英语).
- . Benezit Dictionary of Artists. Oxford University Press http://dx.doi.org/10.1093/benz/9780199773787.article.b00032269. 2011-10-31. 缺少或
|title=为空 (帮助) - Condon, R. . Journal of Gastrointestinal Surgery. 2002-08, 6 (4). ISSN 1091-255X. doi:10.1016/s1091-255x(02)00020-3.
- Knapp, James E.; Carroll, Donald; Lawson, Janet E.; Ernst, Stephen R.; Reed, Lester J.; Hackert, Marvin L. . Protein Science. 2000-01, 9 (1). ISSN 0961-8368. PMC 2144448
 . PMID 10739245. doi:10.1110/ps.9.1.37 (英语).
. PMID 10739245. doi:10.1110/ps.9.1.37 (英语). - Condon, R. . Journal of Gastrointestinal Surgery. 2002-08, 6 (4). ISSN 1091-255X. doi:10.1016/s1091-255x(02)00020-3.
- Voet, Judith G.; Voet, Donald. . Biochemical Education. 2000-05, 28 (3). ISSN 0307-4412. doi:10.1111/j.1539-3429.2000.tb00041.x.
- Fraser, Marie E.; James, Michael N.G.; Bridger, William A.; Wolodko, William T. . Journal of Molecular Biology. 2000-06, 299 (5). doi:10.1006/jmbi.2000.3807 (英语).
- Oyedotun, Kayode S.; Lemire, Bernard D. . Journal of Biological Chemistry. 2004-03, 279 (10). doi:10.1074/jbc.M311876200 (英语).
- Estévez, Marcel; Skarda, Jeremy; Spencer, Josh; Banaszak, Leonard; Weaver, Todd M. . Protein Science. 2002-06, 11 (6). ISSN 0961-8368. PMC 2373640 . PMID 12021453. doi:10.1110/ps.0201502 (英语).
- Lynch, AM; Morton, CC. . Atlas of Genetics and Cytogenetics in Oncology and Haematology. 2011-02, (4). ISSN 1768-3262. doi:10.4267/2042/38351.
- Estévez, Marcel; Skarda, Jeremy; Spencer, Josh; Banaszak, Leonard; Weaver, Todd M. . Protein Science. 2002-06, 11 (6). ISSN 0961-8368. PMC 2373640 . PMID 12021453. doi:10.1110/ps.0201502 (英语).
- Nishiyama, M.; Birktoft, J.J.; Beppu, T. . Journal of Biological Chemistry. 1993-03, 268 (7). ISSN 0021-9258. doi:10.1016/s0021-9258(18)53446-3.
- Owen O, Kalhan S, Hanson R. . J. Biol. Chem. 2002, 277 (34): 30409–12 [2016-08-05]. PMID 12087111. doi:10.1074/jbc.R200006200. (原始内容存档于2008-11-06).
- B. Alberts等著 丁小燕、陳躍磊等譯. . 科學出版社. 2012. ISBN 978-7-03-033015-4.
- Stryer, Lubert. . Fourth. New York: W.H. Freeman and Company. 1995: 509–527, 569–579, 614–616, 638–641, 732–735, 739–748, 770–7731995. ISBN 0 7167 2009 4.