大盗龙类
大盜龍類(Megaraptora)是群肉食性獸腳亞目恐龍,與其他獸腳類間的親緣關係充滿爭議[8][9][10]。其底下的先進物種大盜龍科(Megaraptoridae)以加長的手指爪和大比例的前肢(一般獸腳類前肢比例普遍縮小)聞名。
| 大盗龙类 | |
|---|---|
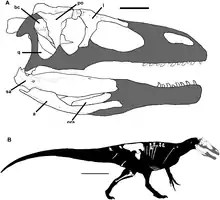 | |
| 巖壁盜龍的頭骨和已知骨骼復原圖 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | 蜥臀目 Saurischia |
| 亚目: | 獸腳亞目 Theropoda |
| 演化支: | 新獸腳類 Neotheropoda |
| 演化支: | 鳥吻類 Averostra |
| 演化支: | 坚尾龙类 Tetanurae |
| 演化支: | 俄里翁龍類 Orionides |
| 演化支: | 鳥獸腳類 Avetheropoda |
| 演化支: | †大盗龙类 Megaraptora Benson, Carrano & Brusatte, 2010 |
| 模式種 | |
| †槊足大盜龍 Megaraptor namunhuaiquii Novas, 1998 | |
| 演化支 | |
目前對大盜龍類充滿未知,因為還沒有發現完整的骨骼。但是大盜龍類仍有許多獨特衍徵。牠們的前肢長而壯碩、進階物種(大盜龍科)的尺骨呈現獨特形狀、前兩根手指加長並長有巨大彎爪、而第三指較小[9]。大盜龍類的頭骨材料十分缺乏,但是一具少年大盜龍標本保存了部分口鼻部,呈現長而纖細,並於2014年被敘述[10]。大盜龍類的腿部骨骼也較細長,類似適應奔跑的虛骨龍類腿部[9]。雖然大盜龍類是身體厚重的獸腳類,牠們的骨骼已大幅氣腔化或充滿氣囊。脊椎、肋骨、骨盆的髂骨氣腔化至獸腳類中非常罕見的程度,也只在新獵龍發現過[8]。其他特徵還包含前後兩側後凹的頸椎、類似美頜龍科的牙齒。
此演化支最早於2010年的研究中命名,並被歸類於新獵龍科之下,這是一群體型輕巧的異特龍超科,與巨大的鯊齒龍科(如南方巨獸龍和鯊齒龍)有親緣關係[8]。一份2013年由費南多·諾瓦斯和同事發表的系統發育學分析反駁這項理論,並提出大盜龍類是從暴龍超科(一群基礎虛骨龍類,包含著名的暴龍)底下分出的演化支[9]。後續的研究修正了Novas的數據和研究方法,支持了第三種可能性:與其他有爭議性的獸腳類如瓜利喬龍位於基礎虛骨龍類,但不屬於暴龍超科[11]。無論牠們真正的分類位置為何,顯然大盜龍類都經歷了似新獵龍型態的異特龍類以及基礎虛骨龍類的大幅度趨同演化[8][9]。
大盜龍類在晚白堊世早期的南美洲達到高度多樣化,尤其在巴塔哥尼亞。而牠們的分布也十分廣泛:布萬獵龍和福井盜龍,最基礎和第二基礎的物種,分別生活在泰國和日本;化石材料也在澳洲常見[12],南方獵龍是該大陸目前已知體型最大的掠食性恐龍[13]。
描述


大盜龍類是一群中至大型獸腳類,從最小型的福井盜龍(4.2公尺)到大型的氣腔龍(9公尺),以及可能的巴哈利亞龍(12公尺)[2]。所有的大盜龍類都來自非常破碎的化石,但許多物種都具有可鑑定的特徵以歸類於此。至少有某些大盜龍類,如氣腔龍和巖壁盜龍,具廣泛的氣腔化骨骼(在肋骨和髂骨最明顯),當中可能有與肺部相接的管道,如同現代鳥類[14]。許多物種具有瘦長的腿骨和長蹠骨表明牠們可能具有擅於奔跑的生活適應[8]。大多數大盜龍類物種屬於大盜龍科,由費南多·諾瓦斯和同事在2013年命名,本科物種的共同特徵包含特化的尺骨和指爪,這些特徵未在基礎物種福井盜龍上發現[9]。
頭骨和牙齒

還沒有發現保存完整頭骨的大盜龍類化石,但許多物種都找到了頭骨碎片。氣腔龍、齒河盜龍、巖壁盜龍保存了頭顱骨的後半部[14],下頜骨則由南方獵龍所得知[13],而一個在2014年描述的大盜龍少年標本保存了部分口鼻部以及頂骨的碎片。牙齒在許多物種都有找到。綜合來說,大盜龍類可以重建為具有長而輕巧的頭骨以及相對細小的牙齒[10]。
根據大盜龍,鼻部尖端的前上頜骨小而長的棒狀分支延伸支撐著外鼻孔,鼻孔本身非常大且加長,類似一些早期暴龍超科(帝龍、原角鼻龍等)。口鼻部亦和鯊齒龍科有相似性,即上頜骨的筆直上緣和矩形的鼻骨。頭骨頂端及眼睛後方的頂骨具有發達的矢狀嵴,如同暴龍超科[10]。另外,頭骨後半部形狀相對簡單,缺乏明顯的頭冠或突起物,但部分物種的淚骨和眶後骨具有皺褶。氣腔龍和巖壁盜龍有著氣腔化的方骨,類似一些異特龍超科(中華盜龍、馬普龍)和暴龍超科[14]。齒骨只有南方獵龍有發現,長而纖巧,第一排牙齒比其他還小(如同暴龍超科)。整個下頜骨只有一個孔洞,就像鯊齒龍類、暴龍超科、似鳥龍類[9]。但下頜骨的後部(根據巖壁盜龍)比暴龍超科更加輕盈。保存下來的腦殼同時與鯊齒龍科和暴龍超科具有相似性[14]。
大盜龍的前上頜牙齒與暴龍超科有許多相似性,小而呈鑿子狀,橫截面為D型[10]。但巖壁盜龍的前上頜牙齒呈犬齒狀,類似非暴龍超科獸腳類。大盜龍類的前上頜齒顯示了物種間的多樣性,牙齒與帶有最小砝硠質鑲嵌的吻部相較之下顯得細小。一些物種,例如齒河盜龍、南方獵龍、大盜龍的牙齒橫截面為8字型且前側完全無鋸齒狀(類似馳龍科、美頜龍科)[10],而牆盜龍僅在齒尖有前鋸齒構造。福井盜龍具有非常側扁、刀刃狀的牙齒(類似鯊齒龍類)並同時在前後側都有鋸齒[14]。
脊椎和肋骨
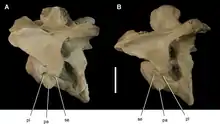
大盜龍類的頸椎明顯的雙側後凹而使牠們在獸腳類中近乎獨特,這表示前側是凸的、後側是凹的。雙側後凹頸椎也是異特龍和蜥腳下目的特徵,可以在不犧牲抗剪力的前提下提高靈活度[15]。此外頸椎神經棘短、中心處肋骨凸面的橫突、和一對大的側凹等特徵也類似於鯊齒龍科。事實上,在大多數大盜龍類的脊椎都有一或多個側凹,並連結到脊椎內部許多氣囊組成的複雜系統。這種網狀內部結構在大盜龍類脊椎(還有少數其他獸腳類)被敘述成像駝峰[8][9]。
末端尾椎沿著下側有道縱脊,類似新獵龍而不像暴龍超科[8]。一對側稜脊向下從橫突延伸至中心(中心突椎板),在橫突下有大凹陷。雖然這樣的稜脊也在背椎和其他獸腳類發現過,大盜龍類獨特在於尾部基幹的中心突椎板發育良好,有時甚至比背椎的發達。只有棘龍科共享此一特徵[14]。發達的稜脊代表著尾部可能厚而肌肉發達[10]。
背肋骨厚且彎曲但中空而在連接背椎處有孔洞。腹肋是寬而堅固的槳狀結構。以上特徵表明大盜龍類和暴龍超科一樣是寬體腔的動物[10]。
前肢
大盜龍類的肱骨呈S形,類似基礎異特龍超科和基礎虛骨龍類。大部分大盜龍類有和異特龍一樣大而健壯的肱骨,但是基礎物種福井盜龍則有較纖細的肱骨[8]。肱骨末端靠近肘部有發育良好的髁和凹槽,類似虛骨龍類尤其是馳龍科。

大盜龍的尺骨有幾項特徵,肘突發達,薄、形狀如刀、沿尺骨縱向延伸,尺骨有長的冠狀結節垂直於肘突,因此尺骨橫截面呈T字型,由向前突起、向外結節向後肘突形成三個尖端。基礎的福井盜龍缺乏此特徵。橈骨與其他獸腳類相比則較為普通[9][16]。
大盜龍類的手部也非常有特色[8][9]。前兩指大而細長,第三指較小。這樣的手指大小差異在某種程度上類似於暴龍超科和其他多種基礎虛骨龍類,但是大盜龍類前肢和手指的發達趨勢則與先進暴龍超科常見的前肢萎縮趨勢相反。大盜龍類保留了第四掌骨(原始獸腳類的第四指)的殘餘部分,這是其他堅尾龍類所缺乏的原始特徵。前兩指有著異常巨大的指爪,大盜龍的第一指爪甚至大於整根尺骨。不像其他獸腳類(如斑龍超科)的大爪,大盜龍的爪橫切面細而呈橢圓形。大盜龍科(非福井盜龍的大盜龍類)爪的平坦面上還有不對稱位置的凹槽,下緣有一道鋒利稜脊。大盜龍類還具有半月形腕骨,類似手盜龍類[16]。
後肢
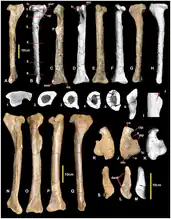
大盗龙类股骨(即腿骨)仅见于南方猎龙和福井盗龙的化石之中,但仅存化石在几个方面与虚骨龙类相似,例如:跗骨发育良好,从股骨干偏移而形成一个深凹。从上面看,跗骨的尺寸使股骨靠近髋关节窝的部分成为矩形。而非虚骨龙类兽脚亚目的跗骨很小,从上面看时股骨呈泪珠状,股骨头略微上翘(如鲨齿龙科和一些虚骨龙类)。在大盗龙类中,由于外侧髁比内侧髁向更远侧突出,从前面看,股骨靠近膝盖的部分不对称。[9]
胫骨也和虚骨龙类相似。这是一根长而纤细的骨头。胫骨外侧髁的前部向下弯曲,类似于新猎龙、长臂猎龙和一些暴龙类。内踝和外踝扩张并相互远离突出(如高等暴龙类和鲨齿龙类。胫骨远端的前表面(靠近脚踝)有一个扁平的小平面,用于连接脚踝的跟骨,情况类似虚骨龙类。小平面内缘被一个脊所界定,这是大盗龙类独有的特征。与异特龙类不同,小平面的上边缘缺乏明显的距骨上柄。位于小平面上的跟骨升突扩展成一个大的梯形骨板,类似于虚骨龙类,但不同于异特龙类的较小的、三角形升突。[9]福井盗龙、南方猎龙和气腔龙在跟骨的外缘上有一个明显的、向前的尖头,前两种恐龙有一个向后突出的额外的尖头。[8]
腓骨也很长,从膝盖开始逐渐变细,就像虚骨龙类一样。它连接到跟骨外缘的一个小面上(虚骨龙类),而不是上边缘的一个大面上(如异特龙类)。腓骨靠近膝盖并面向胫骨的部分有一个宽的凹陷,称为近中窝。第三跖骨连接中趾骨且较为细长,就像虚骨龙类一样。中趾的关节很高,呈滑车状,从下面可以看到一个很深的新月形凹陷。[9]
骨盆

髂骨(髋骨的上半部分)是一根严重气腔化的骨头,充满了气腔并有许多小孔。另一种拥有此特征的已知的大型兽脚类动物是新猎龙。[8]在一些大盗龙类中,前髋臼刃片沿其前缘有一个缺口,类似暴龙类。髋臼前刃和耻骨柄之间的凹陷程度稍低(这个凹陷称为前臼窝)。这种特征存在于各种虚骨龙类、吉兰泰龙的身体结构中,新猎龙也可能拥有此特征。另一方面,髋臼后刃缺少一个大的凹度。在非虚骨龙类的坚尾龙类中,髂骨的这一部分有一个称为短窝的大型凹陷,从髂骨的外表面可以看到。然而,虚骨龙类和大盗龙类有一个小得多的短窝,它只占据了髂骨后缘的一部分,而且大部分对外界观察者来说是隐藏的。[9]
坐骨只存在于巖壁盜龍的化石中。它轻微扩张,类似于鲨齿龙类。[14]耻骨的顶端有一个明显得多的镰刀状扩张,其长度超过骨骼主轴的60%。这种结构被称为耻骨突,在鲨齿龙类和暴龙类的身体结构中也是已知的。耻骨也在与髂骨接触处扩张。左右耻骨没有完全融合在一起,它们沿中线被一个椭圆形的洞分开。[9]
起源和演化
大盜龍類的起源近年來已被確定。古生物學家Phil Bell和Steve Salisbury等人發表的一份研究,關於一種新發現、尚未命名的大盜龍類(被大眾媒體暱稱為「閃電之爪(Lightning Claw)」,可能是盜龍的同物異名)在澳洲閃電嶺西南部田野發現,年代距今1億1000萬年前,揭示了大盜龍類可能起源於澳洲,然後經歷一連串輻射演化擴散至岡瓦納大陸。這份研究也允許了進一步的系統發生學試驗關於大盜龍類究竟來自暴龍超科或鯊齒龍科[12][17]。
分類
早期理論
在大盜龍類演化之於2010年建立以前曾被放置於多個不同的獸腳類群體。大盜龍和福井盜龍於1990年代被發現初期曾被認為是巨大的馳龍科,因為其大型指爪被錯誤鑑定為腳趾爪。但是這些錯誤在重新詳細檢視標本(福井盜龍)和發現新標本(大盜龍)後就得到修正。在2000年代中後期,大盜龍類被認為是基礎堅尾龍類,通常歸類於異特龍超科。Smith等人(2008)提到了澳洲發現像大盜龍的尺骨,並提出大盜龍屬於棘龍科的證據。同一年齒河盜龍被敘述為不尋常的巨型虛骨龍類,與較小的美頜龍科有相似之處。氣腔龍在一年後被敘述成異特龍的親緣物種,而南方獵龍被認為是鯊齒龍科的姊妹群。
歸類在新獵龍科
2000年代後期加入的新資訊促進了幾個主要的基礎堅尾龍類系統發育學的重新分析。Roger Benson、Matt Carrano與Steve Brusatte在2010年的研究發現異特龍超科(或有時稱為肉食龍下目)當中包含了一個主要分支稱為鯊齒龍類,其中又分成鯊齒龍科和新建立的新獵龍科。新獵龍科包含新獵龍、吉蘭泰龍和新命名的大盜龍類。大盜龍類包含大盜龍、福井盜龍、齒河盜龍、氣腔龍、南方獵龍。這幾個物種根據幾個骨骼鑑定特徵被連結在一起,尤其是大幅度的骨骼氣腔化。氣腔龍髂骨的氣腔化相對顯著,而當時新獵龍是唯一也具備此特徵的其他物種。新獵龍科被假想為倖存至最後的異特龍超科,藉由類似虛骨龍類的適應特徵而存活至白堊紀晚期[8]。後續的研究支持此理論,如Carrano、Benson和Sampson(2012)針對堅尾龍類親緣關係的重大研究[18]以及Zanno和Makovicky(2013)敘述新發現獸腳類西雅茨龍將其歸類於大盜龍類。福井盜龍和南方獵龍也持續發現彼此的接近親緣關係,一如氣腔龍與大盜龍之間的關係,齒河盜龍則被視為難以定下正確分類位置。
以下系譜樹根據Benson、Carrano和Brusatte(2010),將大盜龍類歸類於異特龍超科新獵龍科底下的理論[8]。結合Coria & Currie (2016)的研究加入巖壁盜龍(Novas et al., 2013)[14][19]。
| 異特龍超科 Allosauroidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||

歸類在暴龍超科
另一個假設亦在同時期形成:Fernando Novas等人(2012)[20]。Novas和其同僚提出連結新獵龍和大盜龍類的特徵比2010年研究提出的來得普通許多,而與虛骨龍類的趨同演化特徵可能使大盜龍類和虛骨龍類的親緣關係變得有效。此外,他們指出Benson, Carrano, & Brusatte的研究僅列出三個虛骨龍類作為範本。Novas等人的論點在2013年針對巴塔哥尼亞獸腳類的分析中發表,當中將大盜龍類從鯊齒龍類中移除並移至虛骨龍類之下。更精確地說,將大盜龍深置於暴龍超科之下,代表著一支基礎虛骨龍類輻射演化包含著名的暴龍。隨著Novas 等人(2013)將大盜龍類移出新獵龍科,他們建立了新的大盜龍科以包含與大盜龍接近而與基礎福井盜龍較遠的物種。他們還發現幾乎沒有證據能證明吉蘭泰龍、新獵龍或西雅茨龍屬於大盜龍類,而將始暴龍放入大盜龍類。儘管指出大盜龍類與暴龍科的接近親緣關係,Novas等人(2013)註明到大盜龍類形態功能演化方向與暴龍科呈現相反趨勢:暴龍科具有短小手臂、強有力的頭部,而大盜龍類有長有力的手臂、巨型指爪和相對纖弱的頜部[9]。2014年發表的大盜龍少年頭骨標本以與基礎暴龍超科如帝龍頭骨的相似性支持此理論。然而大盜龍類與鯊齒龍類仍有諸多相似之處,其背後之間的分類關係仍未能完全解決[10]。
以下系譜樹支持Novas等人(2013)將大盜龍類歸類於進階暴龍超科的理論。這份研究來自Porfiri等人(2014),其中描述了大盜龍的青少年標本。而瓜里丘龍、巖壁盜龍与特拉塔尼亞龍在這份研究發表時都還沒被描述[10]。
| 鳥獸腳類 Avetheropoda |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||



2016年Novas和他的同僚發表了一份關於大盜龍類手部結構的研究,以幫助解開分類之謎。他們發現大盜龍類缺少許多進階型虛骨龍類(包括五彩冠龍和恐爪龍)手部的關鍵特徵,並具有許多能在異特龍等堅尾龍類發現的原始特徵。至少還有許多特徵支持大盜龍類屬於虛骨龍類[16]。一份由Bell等人2016年發表的關於閃電山脊之爪的研究根據Porfiri等人(2014)分析多項大盜龍類及虛骨龍類的頭骨資訊相較於Benson、Carrano及Brusatte(2010)來支持大盜龍類屬於虛骨龍類[12]。Motta等人(2016)同意,並提到一種破碎的新巴塔哥尼亞獸腳類南盜龍是非大盜龍科的大盜龍類。他們還指出南盜龍與兩種來自非洲、化石被二戰摧毀、神秘的巨型獸腳類三角洲奔龍、巴哈利亞龍之間的相似性。據此他們建議三角洲奔龍和巴哈利亞龍也是基礎大盜龍類,並和南盜龍組成一個獨立的演化支-巴哈利亞龍科[2]。一份由Rolando、Novas及Agnolín在2019年針對巖壁盜龍的重新敘述持續支持大盜龍類位於基礎暴龍超科,以Apesteguia等人(2016)的數據為基礎[21]。
歸類在非暴龍超科的虛骨龍類
2016年,第三个关于大盗龙类分类位置的假设发布,该假设来自Porfiri等人(2014年)对Novas等人研究数据的修订。那一年,塞巴斯汀·阿派斯特圭和他的同事描述了一种不寻常的、全新的兽脚类动物——瓜里丘龙。Porfiri随后在Novas等人的数据集中加入了瓜里丘龙、三角洲奔龙和一些修正,并最终引出了一个有趣的结果:大盗龙类的分类位置反而远离Novas等人最初支持的“基础暴龙类”。而异特龙超科也被列为一个并系群,含新猎龙、一个由吉兰泰龙和瓜里丘龙形成的分支以及大盗龙类,它们在演化关系上逐渐接近传统意义上的虚骨龙类。[6]
Porfifi等人(2018年)的另一项研究通过增加两种大盗龙类,扩展了Apestguía等人(2016年)的研究数据。[11]虽然结果不同,但分析方式实际上与Apestguía等人相同,不同之处仅在于它包含了Apestguía等人分析中未提到的两种大盗龙类。[6]其中一种是巖壁盜龍,在2016年与瓜里丘龙同時被敘述發表。[14]第二种是一属新的大盗龙类,特拉塔尼亞龍。Porfiri等人(2018年)将这两种恐龙列为大盗龙类,将福井盗龙列为最原始的大盗龙类,这一点在Novas等人数据集的所有先前修订中都有发现。然而,大盗龙与吉兰泰龙、瓜里丘龙和暴盗龙类(即传统意义上的“虚骨龙类”)都位于最基础虚骨龙类的一个演化支中。由于新猎龙的分类位置不确定,非虚骨龙类的鸟兽脚类也被分配到了一个大型演化支中。Porfiri等人(2018年)还评论了Motta等人2016年对巴哈利亚龙科的研究,并指出瓜里丘龙也可能属于巴哈利亚龙科,因为它与三角洲奔龙类似。如果是这样的话,那么大盗龙类的前肢形状将具有比以前认为的更多的多样性,如瓜里丘龙的前肢非常小而类似于暴龙。[11]
2018年末,Delcourt与Grillo发表了一项关于于暴龙类的研究。他们重复了Porfiri等人的分析,并在其中增加最近研究的数据。这项研究将新猎龙归入单系的异特龙超科,并将大盗龙类作为与吉兰泰龙和瓜里丘龙演化关系密切的非暴龙类虚骨龙类基群。巖壁盜龍屬也被列为第二种最基础的大盗龙类,排在福井盗龙之前。[22]
以下演化树来源于Delcourt与Grillo2018年的研究。
| 鸟兽脚类 Avetheropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2022年对飔戮龙的描述中,Rolando等人认为大盗龙类处于虚骨龙类内部,并将其列为暴龙超科的姐妹群,与以前那些表明暴龙超科和大盗龙类之间关系密切的研究一致。Rolando等人还注意到大盗龙类中存在两个不同演化支:一个泛围更广,包括除福井盗龙和南方猎龙外的所有大盗龙类(以下写作“演化支A”),另一个泛围更窄,完全由南美大盗龙类构成(以下写作“演化支B”)。以下分支图显示了Rolando等人虚骨龙类系统发育分析的结果:[23]
| 虚骨龙类 Coelurosauria |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
參考資料
- Novas, F., Agnolin, F., Rozadilla, S., Aranciaga-Rolando, A., Brissón-Eli, F., Motta, M., Cerroni, M., Ezcurra, M., Martinelli, A., D'Angelo, J., Álvarez-Herrera, G., Gentil, A., Bogan, S., Chimento, N., García-Marsà, J., Lo Coco, G., Miquel, S., Brito, F., Vera, E., Loinaze, V., Fernandez, M., & Salgado, L. (2019). Paleontological discoveries in the Chorrillo Formation (upper Campanian-lower Maastrichtian, Upper Cretaceous), Santa Cruz Province, Patagonia, Argentina (页面存档备份,存于). Revista del Museo Argentino de Ciencias Naturales, 21(2), 217-293.
- Motta, Matías J.; Aranciaga Rolando, Alexis M.; Rozadilla, Sebastián; Agnolín, Federico E.; Chimento, Nicolás R.; Egli, Federico Brissón; Novas, Fernando E. . New Mexico Museum of Natural History and Science Bulletin. June 2016, 71: 231–253 –ResearchGate.
- Currie, P.J.; Azuma, Y. . J. Paleont. Soc. Korea. 2006, 22 (1): 173–193 –ResearchGate.
- Samathi, A.; Chanthasit, P.; Martin Sander, P. . Acta Palaeontologica Polonica. May 2019. doi:10.4202/app.00540.2018 (英语).
- White, Matt A.; Bell, Phil R.; Poropat, Stephen F.; Pentland, Adele H.; Rigby, Samantha L.; Cook, Alex G.; Sloan, Trish; Elliott, David A. . Royal Society Open Science. 2020, 7 (1): 191462. PMC 7029900
 . PMID 32218963. doi:10.1098/rsos.191462.
. PMID 32218963. doi:10.1098/rsos.191462. - Apesteguía, Sebastián; Smith, Nathan D.; Juárez Valieri, Rubén; Makovicky, Peter J. . PLOS ONE. 2016-07-13, 11 (7): e0157793. ISSN 1932-6203. PMC 4943716 . PMID 27410683. doi:10.1371/journal.pone.0157793.
- Rolando, Alexis; Motta, Matias; Agnolin, Federico; Manabe, Makoto; Tsuihiji, Takanobu; Novas, Fernando. . 2021-12-08 [2022-02-05]. doi:10.21203/rs.3.rs-1152394/v1. (原始内容存档于2022-02-05) (英语).
- Benson, Roger B. J.; Carrano, Matthew T.; Brusatte, Stephen L. . Naturwissenschaften. 2010-01-01, 97 (1): 71–8. ISSN 0028-1042. PMID 19826771. doi:10.1007/s00114-009-0614-x (英语).
- Novas, Fernando E.; Agnolin, Federico L.; Ezcurra, Martin D.; Porfiri, Juan; Canale, Juan I. . Cretaceous Research. October 2013, 45: 174–215 [2020-08-05]. ISSN 0195-6671. doi:10.1016/j.cretres.2013.04.001. (原始内容存档于2018-05-09) (英语).
- Porfiri, Juan D.; Novas, Fernando E.; Calvo, Jorge O.; Agnolín, Federico L.; Ezcurra, Martín D.; Cerda, Ignacio A. . Cretaceous Research. 2014, 51: 35–55 [2020-08-05]. doi:10.1016/j.cretres.2014.04.007. (原始内容存档于2022-04-08) (英语).
- Juan D. Porfiri; Rubén D. Juárez Valieri; Domenica D.D. Santos; Matthew C. Lamanna. . Cretaceous Research. 2018,. in press: 302–319. doi:10.1016/j.cretres.2018.03.014.
- Bell, Phil R.; Cau, Andrea; Fanti, Federico; Smith, Elizabeth T. . Gondwana Research. August 2016, 36: 473–487. doi:10.1016/j.gr.2015.08.004 (英语).
- White, Matt A.; Bell, Phil R.; Cook, Alex G.; Poropat, Stephen F.; Elliott, David A. . PeerJ. 2015-12-15, 3: e1512. ISSN 2167-8359. PMC 4690360 . PMID 26713256. doi:10.7717/peerj.1512 (英语).
- Coria, Rodolfo A.; Currie, Philip J. . PLOS ONE. 2016-07-20, 11 (7): e0157973. ISSN 1932-6203. PMC 4954680 . PMID 27439002. doi:10.1371/journal.pone.0157973.
- Fronimos, John A.; Wilson, Jeffrey A. . Ameghiniana. April 2017, 54 (2): 151–177. ISSN 1851-8044. doi:10.5710/AMGH.12.09.2016.3007.
- Novas, Fernando E.; Aranciaga Rolando, Alexis M.; Agnolín, Federico L. . Memoirs of Museum Victoria. July 2016, 74: 49–61. doi:10.24199/j.mmv.2016.74.05 (英语).
- Pickrell, John. . Blogs. Australian Geographic. Sep 7, 2015 [2020-08-05]. (原始内容存档于2015-09-27).
- Carrano, Matthew T.; Benson, Roger B. J.; Sampson, Scott D. . Journal of Systematic Palaeontology. 2012-06-01, 10 (2): 211–300. ISSN 1477-2019. doi:10.1080/14772019.2011.630927.
- Méndez, Ariel H.; Novas, Fernando E.; Iori, Fabiano V. [First record of Megaraptora (Theropoda, Neovenatoridae) from Brazil]. Comptes Rendus Palevol. May–June 2012, 11 (4): 251–256. doi:10.1016/j.crpv.2011.12.007 (英语).
- Novas, F. E.; Agnolín, F. L.; Ezcurra, M. D.; Canale, J. I.; Porfiri, J. D. . Ameghiniana. 2012, 49 (Suppl): R33 [2020-08-10]. (原始内容存档于2016-03-11).
- Aranciaga Rolando, Alexis M.; Novas, Fernando E.; Agnolín, Federico L. . Cretaceous Research. 1 March 2019,. in press: 104–127. doi:10.1016/j.cretres.2019.02.021.
- Delcourt, Rafael; Grillo, Orlando Nelson. . Palaeogeography, Palaeoclimatology, Palaeoecology. 2018-12-15, 511: 379–387. ISSN 0031-0182. doi:10.1016/j.palaeo.2018.09.003 (英语).
- Rolando, Alexis M. A.; Motta, Matias J.; Agnolín, Federico L.; Manabe, Makoto; Tsuihiji, Takanobu; Novas, Fernando E. . Scientific Reports. 2022-04-26, 12 (1): Article number 6318 [2022-05-25]. doi:10.1038/s41598-022-09272-z . (原始内容存档于2022-04-26).