黑猩猩
黑猩猩(學名:Pan troglodytes)是黑猩猩屬的一種猿類動物,黑猩猩是該屬的兩個物種之一,另一個物種是倭黑猩猩(學名:Pan paniscus)。這兩個物種是當今存在的所有其他動物中基因組成最接近人類的物種,因此引起了科學界和公眾對它們的興趣。[2]
| 黑猩猩 化石时期: | |
|---|---|
 | |
| 東部黑猩猩(學名:Pan troglodytes schweinfurthii,黑猩猩亞種之一)個體,攝於烏干達基巴萊國家公園 | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 哺乳纲 Mammalia |
| 目: | 灵长目 Primates |
| 科: | 人科 Hominidae |
| 属: | 黑猩猩属 Pan |
| 种: | 黑猩猩 P. troglodytes |
| 二名法 | |
| Pan troglodytes (Blumenbach, 1775) | |
| 亚种 | |
| |
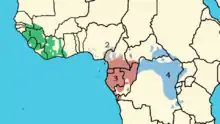 | |
四個亞種的分佈範圍
| |
| 異名 | |
| |
黑猩猩分佈於非洲中部,通常在剛果盆地等熱帶雨林地區中。它們會爬樹,但大部分時間也生活在地面上。它們靠著指關節的外側四肢行走,有時用後腿站立。它的體重略大於倭黑猩猩,但平均小於人類。另一方面,它們的手臂比人類的手臂還要長,體力也大得多。它們是雜食動物,主要以水果為營養來源,也捕食小型哺乳動物。它們非常善於交際,以 20 到 80 個個體為一組生活,並在其間保持著社會等級制度;與倭黑猩猩相反,它們群體中的雄性地位高於雌性。
黑猩猩是目前透過科學研究已知(除人類外)最聰明的動物物種之一。在自然環境中,它們會使用各種工具,例如用來殺死白蟻的棍子和用來敲碎堅果的石頭,有時這些工具以簡單的方式就可以達到預期目的。由於它們個體之間會互相模仿而習得技能,它們各自所屬社群會出現獨特文化與慣習。它們有些個體會在鏡子中認出自己。
它們是瀕危物種。據估計,當前它們在野外生活的個體數目只有十萬到二十萬,而且還在減少。許多威脅嚴重破壞了它們的野生種群,主要是人類的狩獵活動、(人類活動)對它們棲息地森林的破壞,以及明顯經由人類傳播的傳染病。
名稱由來
「Chimpanzee」一詞源於安哥拉地區使用之一種班圖語中的“Kibili-chimpanze”一詞,或可理解為“猿人”,可能字面意思是“類似人的東西”;這種稱呼有時被簡化為“chimp”。
學名Pan troglodytes反映了該物種在科學研究之初的分類地位的混亂,以及神話傳說的影響。troglodytes種小名的意思是“洞穴居民”,這是一個不恰當的名字,因為黑猩猩並不居住在洞穴中。這個名字最初是在 17 世紀由瑞典博物學家卡爾·林奈(Carolus Linnaeus)創造的,作為物種名稱Homo troglodytes(意思是“穴居人”)來表示傳聞存在於非洲的傳說中的人類部落。1776 年,德國博物學家約翰·弗里德里希·布盧門巴赫(Johann Friedrich Blumenbach)將此名稱更改為Simia troglodytes(“穴居猴”)。最後,優先選擇了由德國博物學家洛倫茨·奧肯(Lorenz Oken)於1816年定義的以潘神命名的屬名Pan;潘神在希臘神話中的形象與黑猩猩的相似之處相當微弱。
分類
_and_male_bonobo_(right).jpg.webp)
黑猩猩是黑猩猩屬中的兩個物種之一。另一物種是倭黑猩猩(學名:Pan paniscus),與黑猩猩相似,但體型稍小,身體結構更精緻。
迄今為止,已確定四種黑猩猩亞種,它們之間的區別主要在於它們在非洲的分佈區域:
- 中非亞種(學名:Pan troglodytes troglodytes),主要分佈於加蓬、剛果共和國和剛果民主共和國,以及喀麥隆、中非共和國和赤道幾內亞等政權轄境內。
- 東非亞種(學名:Pan troglodytes schweinfurthii)分佈於坦桑尼亞、盧旺達、布隆迪、烏干達、贊比亞和中非共和國等政權轄境內。
- 西非亞種(學名:Pan troglodytes verus)分佈於塞內加爾、幾內亞、幾內亞比紹、塞拉利昂、科特迪瓦、利比里亞和馬里等政權轄境內。
- 主要分佈於尼日利亞和喀麥隆等政權轄境內的尼日利亞-喀麥隆亞種(學名:Pan troglodytes ellioti 或 Pan troglodytes vellerosus)。
這些亞種之間的外部差異並不顯著,除了西非亞種體型較小的趨勢。遺傳學研究(參見黑猩猩#遺傳學章節)表明,西非亞種和尼日利亞-喀麥隆亞種確實彼此相對隔離(參看異域物種形成),並與其他亞種存在數十萬年的歷史,但中非亞種和尼日利亞-喀麥隆亞種之間沒有顯著的遺傳差異。儘管如此,這些都處於滅絕危險中的亞種的命名法仍用於保存和保護它們的目的。[3]
特徵描述


黑猩猩是無尾靈長目動物。它們的四肢修長而靈活,每條肢體上有五個手指。手指長而發達,並配備了與人類相似的扁平指甲,而不是大多數哺乳動物的爪子。腳的結構類似於手掌,也類似於人的手掌結構,腳趾與其他手指相對,適合抓握樹枝而不適合行走。在黑猩猩的手掌中,手指比人類的手指更長、更強壯,除了拇指較短且不發達。
黑猩猩頭部兩側有貝殼狀耳垂,眼睛大且朝向前方,面部皮膚無毛。它們的牙齒構造與人和其他所有狹鼻小目物種的牙齒構造相同:兩顆門牙、一顆犬齒、兩顆前臼齒和三顆臼齒,共 32 顆牙齒。但與人類(和其他人猿總科動物相似)不同的是,黑猩猩的尖牙又長又尖,尤其是雄性。
與其他人猿總科動物類似,黑猩猩在體型上表現出兩性異形—成年雄性個體體型大而健壯,體重 40至60 公斤,雌性個體體型較小,體重 32至47 公斤。在圈養環境下,黑猩猩有時會因高熱量食物過多而運動量過少而超重,體重可能達到80公斤。當它們四肢站立時,肩部平均高度為82厘米(男性和女性)。當它們用兩隻腳直立時,它們的身高在 100 到 170 厘米之間。與今天生活的所有人猿總科動物一樣,它們的手臂比腿長得多,它們的臂展(從一隻手的指尖到另一隻手的指尖之長度)大約是身高的1.5倍。黑猩猩非常健壯,成年個體體格通常比人類運動員平均而言來得強壯,儘管它們的體重較輕;然而,關於它們比人類強五到十倍的普遍說法是誇大其詞,這源於已被推翻的先前研究。[4]
黑猩猩的身體覆蓋著粗糙的黑色皮毛(有時是非常深的棕色),除了臉、耳朵和裸露的手掌和腳。眉毛和下巴底部的毛髮有時是白色的。幼仔出生時背上有一簇白毛,長大後會變成黑色。幼仔裸露的皮膚顏色非常淺,隨著年齡的增長,不同黑猩猩的顏色往往會不同程度地變深,有時甚至達到全黑皮膚的程度。它們的眼睛通常是棕色的(與大多數非人類哺乳動物相似,“眼白”不是白色),在它們上方是發達的眉脊,尤其是雄性個體,雖然程度不及大猩猩。它們的下巴向前突出,鼻子又小又扁,嘴唇薄而有彈性。
像大多數狹鼻小目物種一樣,它們的臀部襯有堅硬、裸露的皮膚,可以舒適地坐在樹枝或其他堅硬的表面上。發情期的雌性個體屁股腫脹並呈粉紅色,這對雄性具有性吸引力。雄性個體之睾丸大小平均而言相較其他人猿總科物種來得大,是體型大得多的雄性大猩猩的四倍,這可能是因為該物種特有的繁殖方法需要增加精子細胞的產生,群體中的所有雌性都與所有雄性交配。
野生黑猩猩的壽命在 40 到 45 歲之間。
演化
古生物學
黑猩猩的演化史仍然籠罩在迷霧中,因為黑猩猩屬的化石極為稀少。儘管非洲人屬和南方古猿屬的其他原始人類化石相對豐富。與棲息在更開闊或更乾燥地區的物種相比,黑猩猩在熱帶森林中的棲息地可能導致其遺體的保存條件特別不理想。2005 年首次公佈黑猩猩屬的化石:在肯尼亞東非大裂谷東部的一個地質層中發現了許多牙齒大約有50萬年的歷史。牙齒的尺寸相對近似黑猩猩的而非倭黑猩猩,但由於只發現局部,很難確定這些是否確實是已知最早的黑猩猩遺骸。這些化石也可能代表一種已滅絕且未知的黑猩猩屬物種。這一發現證明,該屬的分佈區在更新世時期向今天的分佈區以東數百公里延伸,裂谷並未對黑猩猩屬物種的傳播構成障礙。此外,黑猩猩屬動物和人類似乎在東非有著非常悠久的共存歷史;黑猩猩屬動物種群存在的時間和地點與古代人類物種重合,例如直立人,它們也在同一地層中被發現。[5]
遺傳學
由於化石的稀有性,有關該物種演化的大部分知識都來自其基因研究。由於與人類(在基因等方面)關係密切,黑猩猩是最早被解譯其基因組的哺乳動物之一。黑猩猩基因組計劃於 2003 年 12 月正式啟動,由世界各地的研究機構和大學組成。2005年,計劃團隊發表了全基因組序列的“草稿” 。[6]
已對常見黑猩猩物種的遺傳多樣性進行研究,以便將其與人類物種的遺傳多樣性進行比較。[7][8]出於這些研究的目的,從非洲自然棲息地的黑猩猩身上收集了唾液、血液和糞便樣本,並對它們的部分基因組進行詳盡記錄並相互比較。結果發現,黑猩猩之間的遺傳多樣性明顯大於世界各地人類之間的遺傳多樣性,儘管黑猩猩只生活在非洲的赤道地區。這一發現支持這樣一種信念,即現代人類物種在進化上是年輕的,並且在遺傳上相對同質,並且沒有理由將其劃分為種族或亞種。事實上,這些研究發現,即使在黑猩猩中,也沒有理由將東非亞種與中非亞種分開。然而,分子鐘計算幾項獨立研究表明,幾十萬年前,西非亞種和尼日利亞-喀麥隆亞種彼此分離,並與其他亞種分離。這是一個類似於現代人類從尼安德特人分裂出來的時代。
此外,這些研究表明,黑猩猩和倭黑猩猩在大約 100 萬到 200 萬年前相互分離成兩個物種,儘管它們都生活在赤道非洲,並且只有剛果河將它們的分佈區隔開。結合遺傳學和化石知識,可以推斷出一個近似的系統發生樹(參見人族條目)。
分佈、棲息地與生活方式
黑猩猩的自然分佈範圍位於非洲中部的熱帶地區,位於從塞內加爾轄境延伸到非洲西部大西洋沿岸到坦桑尼亞轄境東部,位於東非大裂谷的南端。在剛果民主共和國轄境,剛果河將南邊的倭黑猩猩和北邊的黑猩猩區隔開來。目前,人類定居和森林砍伐將黑猩猩的分佈限制在該範圍內的偏遠地區。據估計,該物種的大部分野生種群(約 77%)集中在加蓬和剛果之轄境內。據估計,今天的自然種群總數在 100,000 到 200,000 個個體之間。世界各地的動物園裡還有大約 250 個個體,各類研究機構約有 1,200 個個體。
黑猩猩的典型棲息地比其他人科非人類物種更加多樣化。主要包括熱帶雨林,有時是海拔高達3000米的山區,有時是接近海平面的森林。有少量黑猩猩也生活在小型樹林(英語:grove)中,甚至生活在林地和開闊且相對乾燥的疏林草原的混合區域。
儘管黑猩猩個體藉由遺傳獲得優秀攀爬能力,但與狒狒和長臂猿相比,它們相對大的體型限制了其在樹梢上的敏捷移動能力,而且它們大部分時間都在地面上度過。它們是日行性動物,白天覓食,晚上在樹上築巢睡覺。黑猩猩每天晚上都會藉由弄彎幾根相鄰的樹枝並將它們相互組合來建造一個新巢。
在樹頂,黑猩猩以多種方式移動,主要是懸掛在樹枝下並擺動手臂前進。它們長而有力的手臂適合以這種方式運動。他們強大的體力使他們能夠用胳膊和腿從樹枝上爬下,甚至是光滑的樹幹。在地面上,它們主要用四肢行走,依靠指關節的外側—一種黑猩猩和大猩猩特有的行走方式。黑猩猩有時也會用後腿行走很短的距離,例如當它們用手臂攜帶食物時,或者在它們想要讓自己顯得更大時。然而,與人類不同的是,由於他們的腳像手一樣,他們的短腿轉向兩側和背部,因此在能量方面是笨重和浪費的短而相對僵硬,不允許它們完全直立。
在野外沒有觀察到會游泳的黑猩猩,儘管在塞內加爾轄境有黑猩猩在炎熱天氣下會潛入淺水體中以降低體溫。黑猩猩對水的警戒或許可以解釋這樣一個事實,即非洲的河流是物種和亞種分佈範圍之間的天然邊界。動物園有時也會使用這種警告,將它們安置在被充滿水的「護城河」包圍的區域,且已知有黑猩猩在護城河中試圖逃脫而溺水的案例。然而,有些圈養黑猩猩學會了游泳,甚至在游池裡潛水取樂。
黑猩猩是雜食動物。它們主要以水果為食,其中與無花果榕相似的各種熱帶榕屬植物佔有重要地位。它們還吃樹葉、葉芽、莖、花序、樹皮、樹脂,有時甚至吃一點泥土,這些顯然是礦物質的來源。它們以動物為食,捕捉無脊椎動物,從鳥巢中拿取鳥蛋,從蜂巢中取出蜂蜜,並獵殺小型哺乳動物。它們可能會花很多時間將白蟻趕出其巢穴。野生黑猩猩被觀察到捕食較小的猴子,主要是紅疣猴屬(Red colobus)動物,還有藪羚、麂羚和疣豬幼仔。動物最多佔野生黑猩猩飲食的 3%,但非常被它們重視,具有社會重要性。
黑猩猩除人類外幾乎沒有天敵。非洲豹能夠爬樹,且眾所周知有捕食有尾猿猴動物的傾向,因此據推斷,它是大部分分佈地區發現的黑猩猩的主要捕食者,但直接證據極為罕見。白天成群結隊時,黑猩猩會聯合起來對抗非洲豹等動物,甚至扔樹枝和石頭攻擊之,但有理由認為,在夜間,單隻黑猩猩會更容易受到攻擊。在大多數黑猩猩的棲息地都沒有發現獅子,因為獅子的典型棲息地是開闊的疏林草原,而不是大多黑猩猩所在的熱帶雨林。
行為
黑猩猩是極具群居傾向的動物。它們以 20 到 80 個個體為一組生活,規模比任何其他人科非人類物種都相較大。每個群體的領地在森林地區為 5至40 平方公里,但在熱帶草原地區(食物來源密度低得多)可能達 560 平方公里。
大群經常分裂成更小和靈活的群體,為了尋找食物,這些群體可能在一天內彼此分開,甚至長達幾天。由於它們的食物來源,尤其是時令水果,經常供應不穩定,因此黑猩猩一天中的大部分時間都被迫尋找它們,同時還要不斷巡邏它們的大片領地。一棵結著成熟果實的樹並不總是能為幾十個個體提供足夠的食物,因此黑猩猩更願意在一個小群體中尋找食物,而不是與該群體的所有其他成員爭奪食物。當成群的黑猩猩在森林地面上巡邏時,它們的行動通常是安靜而有紀律的,排在由年長且經驗豐富的黑猩猩領導的後方縱隊中,這些黑猩猩警告跟隨它們的人可能存在的危險。
群體中的個體地位
一群黑猩猩由最成熟和最強大的雄性個體統治,其中有一個社會等級,從最高級(“雄性領袖”)到最低級。雌性的地位低於雄性(與倭黑猩猩相反),但它們之間也存在社會等級。
占主導地位的雄性小心翼翼地確立並維持自己在等級中的地位,主要是藉由露出尖牙等各種權力「展示」行為,又如敲擊樹幹、撼搖樹枝和投擲大石塊。這種表演很少演變成真正的暴力戰鬥,因為群體成員認識所有其他成員並清楚地記住其身份。地位低下的雄性和雌性通常會以順從和安撫的姿態回應占主導地位的雄性的表現,以確認其地位。甚至雄性之間爭奪地位的比賽也可能被視為炫耀性比賽,雙方都試圖在不戰鬥的情況下以實力打動對方,直到其中一方以屈服的姿態認輸。然而,極少數情況下,這種鬥爭是在戰鬥中決定的,戰鬥是通過毆打、踢和咬傷進行的,並可能以重傷甚至死亡告終。
許多研究人員,例如珍·古德(Jane Goodall)和其團隊以及弗蘭斯·德瓦爾(France de Waal),多年來一直在紀錄黑猩猩群體的地位鬥爭和變化。他們注意到,獲勝者往往不是最強壯、最具攻擊性的雄性,而是那些擅長與其他雄性和雌性建立友誼、招募盟友和建立“聯盟”的雄性。德瓦爾將黑猩猩群體中的等級鬥爭定義為「政治」。與智力較低的動物物種群體的情況不同,黑猩猩統治者不能對他的臣民行事過於專橫,即使他比任何其他雄性都更有權力,因為他很快就會被許多群體成員聯合起來反對。一個典型的雄性首領之所以統治,要歸功於他與其他雄性(有時是其兄弟)建立的友誼,並與之分享食物和接觸受歡迎的雌性等好處。
雌性通常不會出現群體地位之「展演」行為,而且它們之間的等級比雄性之間的等級更不容易發生逆轉。它主要由經驗和資歷決定,通常由最年長的女性領導,有時女性擁有數名高地位雄性後代。然而,即使在雌性之間,關於地位的衝突也很少會演變成打鬥,儘管沒有雄性之間的衝突那麼危險。
像所有其他猴子和猿類動物一樣,黑猩猩會花費大量時間“梳理”其他黑猩猩的皮毛(參見社交梳理)。這種行為是緩解群體成員之間緊張關係和加強社會聯繫的主要手段。占主導地位的雄性會與其他雄性甚至是雌性結成聯盟,而地位低下者則會為年長者梳理,以示和解或請求恩惠。同樣,雌性梳理其子代個體,而年輕個體為其母親梳理,也與其他年輕個體互相梳理。
群體之間的衝突
與它們在群體中的社交行為相反,黑猩猩群體之間經常表現出明顯敵意。[9]
群體中的雄性會組隊巡邏群體領地邊界,有時也會有少數雌性伴隨。德瓦爾指出,在位在荷蘭阿納姆的Royal Burgers'動物園,即使是從未見過其他群體的雄性,也會每天例行地在它們居住的人工島沿岸周圍巡邏。在野外,這種巡邏有時會升級為有組織的“突襲”,深入到鄰近群體的領地。巡邏和突襲的參與者走在後方縱隊中,同時保持緊張和紀律性的沉默,偶爾停下來傾聽,希望能突襲伏擊落單個體或來自其他群體的小群體,並儘量不陷入反伏擊。這種伏擊有時會以重傷和死亡告終,尤其是當一方設法建立對另一方的數量優勢時。在至少兩個記錄在案的案例中,在位在坦尚尼亞轄境內的貢貝溪國家公園等地,大群黑猩猩在持續數年的一系列襲擊中一個接一個地消滅了較小的鄰近群體的成員,並最終佔領了它們的領地。[10]
在野外,只有年輕的雌性有時會離開它們的群體,並被安全地接納到另一個群體中。雄性不會被接受到另一個群體中,但是一群雄性和雌性可能會從一個大群體中分離,以便在未被佔領的土地上建立一個新的群體。
即使在圈養條件下,也很難使黑猩猩群體接受未知的成年個體加入它們。德瓦爾報告了一個案例,他們試圖說服研究機構中的一群雌性接受兩名雄性。雌性的反應如此猛烈,以至於它們不得不將其中一隻雄性分開以保護其性命。然而,兩隻雌性梳理了第二隻雄性,並保護他免受其他雌性的傷害,直到他被群體接納;向前追溯,這兩個雌性個體在早些年前在另一研究機構認識了這個雄性個體。
溝通

已知有數十種手勢、面部表情和聲音用於它們之間的溝通[11]。其中,以下是特別知名的:
- Pant-hoot:這是黑猩猩最響亮的叫聲。它以一系列快速的喘息開始,最終以尖銳的尖叫聲告終。這種叫聲表示非常興奮,通常在群體中進行,似乎主要用作在其他黑猩猩個體和群體面前展示存在感。
- Waah吠:黑猩猩與另一被攻擊個體發生接觸時發出的警告性叫聲,可能被用作對攻擊者的威脅。
- Tonal-Barks:發現蛇時發出的特有警告叫聲。
- Ho:一種高亢的叫聲,用作溫和的警告,表示尷尬或輕微的焦慮。
- 咕嚕咕嚕聲(英語:pant-grunt):一連串快速的咕嚕聲,是群體中低位階個體向高位階者發出的和解和征服的呼喚。
- 笑聲:與人類的笑聲相似但不完全相同。它由一系列無聲的喘息組成,嘴巴張開,但嘴唇隱藏著牙齒。這是黑猩猩幼仔對搔癢的典型反應,它們通常對搔癢表現出非常積極的情緒反應。
- 微笑:張嘴露出牙齒,表示恐懼而非愉悅。這種反應在參加人類娛樂表演的圈養黑猩猩中很常見,之所以這樣命名是因為人類觀眾經常將其誤認為等同於人類的微笑。
許多黑猩猩的手勢容易被人類識別。例如,向上張開手掌請求給予物件(經常是食物),並拉著其他黑猩猩或人類的手臂請求跟隨它們。它們藉由擁抱和親吻作為親密和和解的象徵(但不作為性行為相關表現),並藉著肢體碰觸及輕柔撫摸來緩和彼此身心狀態。黑猩猩會因缺乏關注而感到沮喪,如正在接受哺乳的幼仔會像人類幼兒一樣,藉由哭鬧表現對於不被理會的不滿情緒。但是,與人類不同,它們通常不會用手指來引起注意,並且它們需要時間來理解人類藉由手指表達的意思。
狩獵與物種間關係
黑猩猩的狩獵活動主要由雄性進行,通常成群結隊。首選的獵物是小型猴子,通常是紅疣猴屬(英語:Red colobus)動物,它們比黑猩猩爬得更快,能夠逃到更細的樹枝上,並從一棵樹跳到另一棵樹。因此,必須先將它們圍困在一棵孤立的樹頂上,然後才能進行捕捉活動,且通常被捕者是體型較小且缺乏生存經驗的幼仔。在位在坦尚尼亞轄境內的貢貝溪國家公園,某些雄性黑猩猩表現出了獨自捕獵的天賦,也許是因為那裡的森林不那麼高大茂密,更容易在孤立的樹上捕捉獵物。研究人員對黑猩猩之間在狩獵中的合作程度仍未達成共識。西非亞種的成員顯然更有組織性,研究確定戰術角色是有計劃和劃分的,比如隱藏起來等待獵物的“潛伏者”,負責擊殺獵物的“刺激者”,以及負責引導有經驗的人引導方向的“狙擊者”的伏擊者。但在中非和東非亞種的大多數群落中,狩獵似乎更具機會主義色彩,組織性更差,隊伍中的每個成員都選擇自己的行動方式。捕獲的獵物藉由撕咬、(藉由四肢)撕成碎片或將之猛擊樹幹而殺死。[10]
黑猩猩十分愛好野生動物的肉,捕獲獵物後會展開爭奪。捕捉到獵物的雌性通常會被迫立即將其交給更強壯的雄性,這解釋了它們在狩獵活動中的參與度較低。雄性通常會優先獲得它們捕獲的獵物,即使是地位較高的雄性,但會立即被其他群體成員包圍,乞求分享,並且通常會至少讓出其中一些。西非亞種傾向於將獵物主要分配給其他狩獵參與者。但即使是沒有參與的雌性,如果足夠堅持,也幾乎總能贏得一些份額,尤其是在發情期。將獵物分配給雄性和雌性的雄性往後可能會在爭取高階地位的鬥爭中獲得受贈者的支持,而雌性可能更願意與與它們分享肉的雄性交配。[10]
珍·古德聲稱,貢貝溪國家公園的黑猩猩和狒狒屬動物之間的關係十分複雜;它們通常共享相同的棲息地,並圍繞相同的食物來源聚集在一起。黑猩猩有時會獵殺狒狒幼仔作為食物,就像它們獵殺其他猴類一樣,但它們會尊重成年狒狒,尤其是雄性狒狒,它們體型幾乎一樣大且強壯,有更長且鋒利的獠牙。黑猩猩會對狒狒發出的瘋狂警告呼喊聲做出反應,反之亦然。黑猩猩幼仔和狒狒幼仔可能一起玩耍,甚至在一段時間內結成友誼。兩物種的雄性,尤其是年輕的雄性在對性活動產生興趣時,會對對方物種雌性的與發情直接相關的身體部位腫脹現象做出反應,並被觀察到(跨物種)交配行為之嘗試,至少在一個案例中因雌性的配合成功完成。
在圈養環境中,年輕的黑猩猩通常容易與家犬、家貓和動物園裡的許多其他物種個體發生積極互動關係。它們會學習照顧其他物種的幼仔,甚至用奶瓶餵養它們。
交配行為
雄性9歲進入青春期,16歲性成熟。在此期間,它們確立了高於群體中雌性(包括它們的母親)的地位,並開始對成年黑猩猩的性活動表現出興趣。雌性平均在 10 歲時第一次交配,但它們在第一次發情週期通常不育。在此期間,它們有時會離開自己的群體並加入另一群體,因為它們的性吸引力保護他們免受來自不熟識它們的雄性的攻擊。它們第一次分娩的平均年齡為 13 或 14 歲。
黑猩猩一年中沒有固定的繁殖季節,儘管雌性黑猩猩更有可能在食物豐富的時候交配。他們的排卵週期約為36天。發情期在排卵期間持續約10天,其特徵是雌性的生殖器等部位腫脹,從而吸引雄性。在發情期間,雌性願意與群體中的大部分或所有成年雄性交配。珍·古德和其他野生黑猩猩研究人員有時會觀察到雄性會聚集在一隻受歡迎的雌性周圍,平靜而有耐心地等候與之交配的機會。占主導地位的雄性(或另一年長雄性個體)可能會試圖將「已婚」雌性隔離開來,以防止該雌性與其他雄性交配,但這種情形通常不會發生在該雄性身上。然而,雌性往往更頻繁地與更年長的雄性交配,並與與它分享食物的雄性交配,因此這些雄性產生後代的機會更多。[12]
由於上述之繁殖方式,群體成員無法知道後代的父親是誰,雄性也不會表現出育幼的傾向和行為。弗蘭斯·德瓦爾認為,這種方法是一種經由演化產生的「策略」,可以降低殺死幼仔(英語:Infanticide)行為的可能性。在其他一些社會動物物種中,包括猴子物種,新的優勢雄性有時會殺死先前雄性的幼仔,以使雌性與它們建立聯繫並交配,但在黑猩猩中,這樣做的雄性可能會殺死自己的後代。在黑猩猩中確實有觀察到幼仔被殺的案例,但這種情況非常罕見。
在懷孕期間和產仔後的三到五年內,雌性在照料和哺育後代期間不交配,也不進行性活動。在這一點上與動物界的大多數雌性相似,不同於雌性人類和雌性倭黑猩猩,後者甚至在排卵期之外也在性行為方面保持活躍。
育幼


.jpg.webp)


雌性黑猩猩的妊娠持續八個月。通常只有一隻幼仔出生,但與人類一樣,雙胞胎的情況也存在。然而,由於雌性黑猩猩獨自撫養幼仔,攜帶雙胞胎和照顧它們非常困難,野生黑猩猩只有極少數例記錄顯示雙胞胎存活到成年。
與大多數其他種類的猴子和猿類一樣,幼仔最初靠在母親的肚子上,緊貼著其皮毛,後來逐漸開始騎在母親背上。在黑猩猩身上,被抱在肚子裡的階段持續五到六個月,前兩個月幼仔是完全無力的,母親必須一直扶著它,因為這個階段的抓握反射是還不夠強。在大約六個月大的時候,幼仔可以騎在母親的骨盆上,同時扶著母親的背部。直到一歲,幼仔一直與母親保持接觸;兩歲時,它會在獨立測量和坐下時遠離她約五米。幼仔在 4 至 6 歲時斷奶,但繼續陪伴並接受母親的幫助尋找食物、指導和保護,直到 9 歲左右成年。
當雌性個體停止母乳喂養時,它能夠再次交配並懷孕,因此手足間的年齡差異通常為四到七歲。儘管只有三年相差的罕見案例已被記錄下來,但這種相差給母親帶來了很大的負擔,它必須同時背著一隻年幼的幼仔和一隻年長的幼仔。黑猩猩幼仔有時會在手足出生時表現出嫉妒,從而引起母親的注意,但在很多情況下,幼仔對手足很感興趣,甚至會幫助母親撫養它們。尤其是雌性,可能會從照看手足的過程中學到很多關於育幼的知識,這些知識在它們自己育幼的時候會產生極大助益。
許多黑猩猩,無論是雄性和雌性,即使在長大後仍與它們的母親和手足保持密切關係。由地位高的母親養大的黑猩猩通常表現出更多的自信,成年後它們在獲得高地位方面具有優勢,尤其是當它們與手足合作時。在貢貝溪國家公園的一個群體中,一個年長的雌性個體至少生了五個後代,其中有兩個雄性後代成為群體領導和副領導,還有一個雌性後代繼承了它的年長雌性地位。一個雌性後代總共生了九個後代,其中七個存活到成年,其中三個是雄性,曾一度成為群體領導。
成年雄性通常不參與育幼活動,但通常會避免傷害它們,並對它們表現出比對群體中的成年成員更寬容的態度。華特迪士尼公司出品的自然類紀錄片《黑猩猩的世界》(2012年)記錄了象牙海岸之塔伊國家公園發生的一個真實且不尋常的案例,群體裡的年長雄性收養了一隻喪母的幼仔,甚至還經常將它背在背上。古道爾記錄了貢貝溪國家公園中群體中的 13 個喪母幼仔,其中七個被成年雌性手足收養,兩個被成年雄性手足收養。
認知能力
黑猩猩是人們迄今已知僅次於人類的最聰明動物物種的候選者。倭黑猩猩的智力可能與黑猩猩相似,但由於個體數較少等原因,相關研究較少。很少有其他動物種類能與某些地區的黑猩猩的表現相媲美,例如許多海豚能在鏡子中認出自己,一隻名叫Alex的非洲灰鸚鵡表現出高度語言和邏輯能力,新喀鴉使用工具甚至改造之。但只有黑猩猩在所有這些領域,甚至在其他領域都表現出色。
使用工具
黑猩猩使用各種不同的工具。它們謹慎地選擇這些工具,甚至以簡單的方式改造之以適應所需用途。在野外中被充分研究的大多數社群中,記錄了 10 到 20 種不同的技能,但由於不同社群之間的技能沒有完全重疊,記錄的工具使用技能的總數比這更多,達到幾十種。[13]其中尤為出名的是:
- 用於捕捉白蟻的棍子:有黑猩猩將細棍子穿入白蟻蟻丘的洞穴開口,它首先用手指將其擴大。白蟻用它們的口器掛在棍子上攻擊棍子,黑猩猩小心地取下棍子並舔食棍子上的白蟻。為此,黑猩猩有時會剪下小樹枝,撕下樹葉以備使用。1960 年,珍·古德在貢貝溪國家公園發現這種行為。試圖模仿這種行為的人類觀察員指出,要正確執行這種行為需要高超的技巧,而且白蟻吃起來很美味。
- 用石頭敲碎堅果:可能是在野生黑猩猩中發現的最複雜的工具使用方法,因為它需要結合一塊扁平且相對較大的石頭,用作放置堅果的平面,還有一塊較小的石頭,作為敲擊用之工具。即使在這種情況下也需要技巧,以便在不壓碎果核的情況下打破堅果外殼。黑猩猩蒐集合適形狀的石塊作為敲擊底座,並將之搬運到特定的擊碎堅果場地。考古學家在象牙海岸發現了藉由放射性碳定年法判定距今4300年的堅果擊碎場地遺址,並由此發展“靈長目動物考古學”。[14]
- 用於獲取蜂蜜的棍子:它可能包括三到五根不同形狀的棍子,以適應不同的功能,一個接一個地執行相同的目的—從蜂巢中獲取蜂蜜。細而鋒利的棍子用於在巢穴中穿孔,粗而重的棍子用於擴大開口,樹皮片用作勺子以裝盛蜂蜜。[15]
- 打獵用的鋒利棍子:這種棍子被用作長槍,用來殺死躲在樹幹裡的小動物。黑猩猩可以藉著將樹枝折斷至正確的長度、去除頂部的樹皮並用牙齒磨尖來改造樹枝。將鋒利的一端穿入錐子並用力擊打以殺死受害者,然後折斷錐子的側面以取出屍體。在塞內加爾進行研究的靈長目動物學者 Jill D. Pruetz 報導了這種行為,目的是獵殺嬰猴。[16]與主要由雄性進行的狩獵不同,在該案例中使用鋒利的棍子進行的狩獵主要由年輕和青春期的雌性進行。[17]
- 樹枝和石頭作為武器:雄性黑猩猩經常在展示時扔樹枝和石頭,但在實際戰鬥中幾乎從不使用它們作為武器,也許是因為這種使用限制了它們四肢奔跑和攀爬。
- 築巢過夜:黑猩猩將幾根樹枝彎曲,並將它們相互組合,形成一個直徑超過一米的表面。它們用柔軟的樹葉把巢排成一行,有時甚至用特別柔軟的樹葉在它們的頭下做一個枕頭。所有在野外被研究過的現存黑猩猩都會築巢,它們在同一個巢穴中的睡眠時間不會超過一晚,因此它們每天晚上都必須築一個新巢。
黑猩猩通常藉由模仿其他黑猩猩的行為來學習使用工具的技能,以及許多其他習俗和技能。幼仔往往會仔細觀察母親的行為,或者有時觀察它們的手足和其他群體成員,並花費數小時試圖模仿它們。因此,每個社群的“傳統”和“文化”特徵都是自然形成的,其習俗與其他社群的習俗不同。[18]例如,在象牙海岸的塔伊國家公園,黑猩猩徒手捕捉白蟻;在貢貝溪國家公園的黑猩猩則如上述使用工具。在塔伊國家公園有用石頭敲擊堅果的群體,鄰近的群體中有以樹根為基座、以木棍為敲擊工具的傳統。不同傳統之間差異的原因仍在研究中。[19]
有時黑猩猩會自己發明工具的新用途。在貢貝溪國家公園的群體中,其中一個雄性個體發現,用它從珍·古德駐紮營地取得的空塑膠容器來「展演」會發出很大的噪音,比用石頭和樹枝來展演更能在觀眾中留下深刻印象。它在跑動時練習在身前滾動兩個塑膠容器,顯然它提前計劃了展演行為。這種使用並沒有成為一種傳統,因為研究人員隨後開始隱藏塑膠容器,但在這個階段,雄性已經成為了群體的領導者,並保持了這個位置好幾年。
在圈養環境中,黑猩猩學會許多使用人類工具的技能,既有主動訓練,也有自發模仿,如泡茶、用器具進食、在紙上用顏料作畫、吸菸、騎乘自行車和各種交通工具等。在動物園裡,黑猩猩母親們學會了用奶瓶餵養幼仔。
符號語言和抽象思考
圈養黑猩猩參加了各種旨在教它們使用自然語言或人工語言進行交流的實驗。它們不能說話是因為喉嚨的解剖結構不適合,但可以使用許多不需說話的符號語言中的單詞。黑猩猩華秀是學習手語的先驅,其他幾個個體也緊隨其後。它們中的大多數掌握了一百多個符號的詞彙,並藉之理解給它們的指示,提出簡單的要求,在某些情況下甚至發明了自己的單詞組合。然而,這些黑猩猩並沒有根據句法規則自發地將單詞添加到句子中,像幼年人類個體那樣。有些黑猩猩學會使用彩色塑膠製符號象徵物進行溝通,或者學會使用特殊鍵盤在計算機螢幕上鍵入的符號進行溝通,它們確實遵從句法規則構造並理解了十個或更多符號的句子,但它們並不是自發地這樣做,而是被人類刻意訓練的結果。語言學家之間存在爭議,這些成就在多大程度上可以被視為符號語言,或者它們是否只是動物行為機制的非常詳盡的例子,例如操作性條件反射。
圈養的黑猩猩展示了基本的數學技能,例如對小數進行除法和加法。例如,他們始終能夠從兩個托盤中選擇包含較大巧克力塊的那個,即使這些塊被分成不同數量的組。在京都大學進行的實驗中,一個雌性黑猩猩學會了識別從 1 到 9 的數字,按其順序排列之,並將每個數字與任何物體的適當數量聯繫起來。在學會這個之後,它被介紹認識數字零,它學會了以正確的順序排列之並將其與沒有任何物體相關聯,儘管並不完美。[20]
視覺記憶和戰略思考
在京都大學進行的實驗中,還發現黑猩猩能夠比大多數人類更好地執行某些視覺記憶任務。在雌性“E”和其雄性幼仔“Iomu”學會按順序敲擊觸控螢幕上顯示的 1 到 19 數字後,研究者使任務變得更加困難—數字在螢幕上不定處閃爍,並在第一個數字被敲擊時隱藏在方塊中,黑猩猩仍能以正確的順序敲擊方塊。在嘗試過這項任務的人類中,沒有人的表現比得上黑猩猩。
在另一項任務中,它們在一種“捉迷藏”的電腦遊戲中競爭,每個玩家都需要藉由點擊觸控螢幕,在左右兩側之間進行選擇,如果“捕手”獲勝它能夠選擇“隱藏”選擇的一側,而如果它設法選擇與捕手選擇的相反一側,則隱藏獲勝。在進行這樣一輪又一輪的博弈時,根據前一輪的結果進行選擇時,根據博弈論的數學理論,選擇的最優策略是由一個納什均衡決定。結果發現,它們對戰時的策略非常接近這個最優策略,而人類玩家在同一局對戰時策略就不太好,平均與最優策略相距甚遠距離是黑猩猩的 6 倍。此外,當獲勝者的獎勵(以黑猩猩的食物和人類的金錢獎勵的形式)增加時,黑猩猩的策略按照博弈論的預期以比策略更快更好的方式改變的人類玩家。京都大學的研究人員假設,黑猩猩比人類更善於記住前幾輪的結果,也許它們比人類更擅長玩零和遊戲,因為它們不像人類那樣願意相互合作。[21]
在靈長目動物學家弗蘭斯·德瓦爾的參與下,研究人員找到了一種在最後通牒賽局中測試黑猩猩的方法:在有獎品(例如一些香蕉)的情況下,實驗者讓“出價”的黑猩猩可以選擇兩種不同的代幣,一種代表獎品的平均分配,另一種代表不平等分配其中投標著收到大部分香蕉。提議者拿走自己選擇的代幣後,可以將其傳遞給“響應者”黑猩猩。如果響應者將代幣返回給實驗者,這被解釋為接受提議,並且實驗者根據選擇的代幣將香蕉平均或不平均地分配給黑猩猩。但是如果響應者拒絕了這個提議,它就會扔掉代幣或者不把它還給實驗者,在這種情況下,沒有一隻黑猩猩會收到香蕉。在這個實驗中發現,黑猩猩更喜歡平等和“公平”地分配獎品,這與人類兒童以相同參與方法的結果相似,也與大多數成年人為金錢獎品而參與時相似,但與根據博弈論而來的理性策略相反。[22]
自我意識與心智理論
黑猩猩學會藉由視覺識別自己,一些研究人員認為這種能力是自我意識的證據。[23]在圈養環境中,習慣於照鏡子的黑猩猩用它們來梳理自己和檢查牙齒。它們很容易通過自我識別測試,在他們不知情的情況下,在其臉上畫了一種顏色,當它們在鏡子中看到自己的倒影時,它們的反應是觸摸臉上的色點而不是倒影。目前還沒有猴科和闊鼻小目動物通過這個測試,就連其他種類的猿類也難以通過。
各方觀察者和研究人員報導了一些案例,在這些案例中,黑猩猩的行為方式暗示著一種精神生活、一種儀式或一種審美意識,但很難證明確實如此。珍·古德報導了幾個案例,在這些案例中,貢貝溪國家公園的群體對雷暴和突如其來的暴雨做出反應,雄性個體反覆出現「展演」行為,而雌性則坐著觀看。珍·古德認為這些表演是針對雨的,稱它們為“雨舞”,並暗示這是萬物有靈論的一種原始形式。後來發現,野生黑猩猩有時會對「繁忙」流動的瀑布做出類似反應,而研究塞內加爾另一黑猩猩群體的靈長目動物學家 Jill D. Pruetz 也報導了對森林火災的類似反應。在貢貝溪國家公園,黑猩猩幼仔通常被抱在懷裡,「梳理」著石頭和棍子,模仿育幼行為,類似人類孩童玩洋娃娃那樣。動物行為學學者 Adriaan Kortlandt 報導了“一隻黑猩猩連續 15 分鐘凝視著特別美麗的日落,觀察不斷變化的顏色,直到它變得太暗而看不見”,珍·古德也報導了類似的行為。黑猩猩有時會在樹幹上敲擊以在「展演」期間發出噪音,但在貢貝溪國家公園,記錄到一隻雄性黑猩猩過去常常敲擊許多不同的物體來取樂,顯然是因為它喜歡發出的聲音。
研究人員仍不同意黑猩猩在多大程度上擁有心智理論,即它們是否將意圖、信念和精神狀態歸因於其他生物(其他黑猩猩或人類)。檢測這種能力的各種心理實驗經常在人類兒童中進行,並表明他們在 5 歲左右時就獲得了這種能力。其中一些實驗(或類似的實驗)是在黑猩猩身上進行的。在一個這樣的實驗中,黑猩猩觀察一個人執行一個異常簡單的動作,例如藉由推動他的膝蓋來打開電子開關。實驗組的黑猩猩也被展示了這種不尋常舉止的可能“原因”——這個人的手正忙著拿著一些東西,而對照組的黑猩猩沒有被展示任何原因。然後給黑猩猩模仿人類的機會,實驗組的黑猩猩正常地用手打開開關,而對照組的黑猩猩準確地模仿了這個不尋常的動作。從這樣的結果中得到公認的結論是,實驗組的黑猩猩將人類的意圖解釋為以通常的方式打開開關。
大多研究者從多次此類實驗的結果中得出的結論是,黑猩猩確實知道如何以與自己相似的信念和意圖對待其他生物,因此它們也能產生與人類相似的同理心。然而,它們似乎很難將不同於它們自己的信仰和意圖歸因於其他生物。也就是說,黑猩猩將無法通過 Simon Baron-Cohen 的“Sally–Anne 測試”。
然而,黑猩猩仍能在某些類似於自然情況的情況下表現出錯誤信念的歸因。這種典型的情況是偷竊另一個生物。例如,當黑猩猩可以選擇從實驗者附近的兩個食物獎品中選擇一個時,它們會更願意抓住被隔板擋在實驗者視線之外的獎品。它們知道如何做到這一點,即使實驗者本人被隱藏起來並且它們看不到他正在看的地方,但它們可以從隔板的位置推斷出他能看到和不能看到的兩個獎品中的哪一個。這是一個非常簡單的欺詐示例,基於將缺乏知識歸因於欺詐的受害者。[24][25]
人類與黑猩猩
人類對黑猩猩的研究歷程
中非原住民對黑猩猩十分了解,直到今天,他們其中一些人仍將黑猩猩幼仔當作寵物飼養。然而,在西方文化中,黑猩猩和其他人科非人類物種在古代和近代初期並不為人所知。歐洲不時有人科非人類物種和類人生物存在的傳言,主要來自船隻失事被迫在非洲海岸逗留一段時間的水手,但這些傳言包含非常多虛構元素,以致很難確定它們在多大程度上基於真實觀察或關於傳說和小說。1641年荷蘭醫師尼古拉斯·杜尔所描繪的生物無疑是一隻猿猴,很可能是由從熱帶地區返鄉的水手攜帶,但不確定是黑猩猩還是紅毛猩猩。
最初的可靠科學描述出現在17世紀末,由英國醫師和解剖學家爱德华·泰森完成,有時被認為是比較解剖學領域的鼻祖。這個個體是在現在被稱作安哥拉的地區捕獲的,但沒有在乘船前往英國的旅程中倖存下來,但泰森在其尚存活時觀察之,死後為之進行解剖。他稱這個生物為“Pygmie”,意為矮人,但從詳細的描述和圖畫來看,毫無疑問它是一隻年輕的黑猩猩,它的骨架至今仍陳列在倫敦自然史博物館。泰森表示它不是人類,但它更像人類而不是有尾巴的猴子,從而落入今天公認的生物分類學的觀點。泰森認為它在「存在鎖鏈」中處於動物和人類之間的中間位置;演化論的第一個版本在泰森死後約一個世紀被提出。
1738 年,倫敦一家報紙宣布一艘商船從安哥拉運來“一種極其醜陋的動物……叫做chimpanzee”。這可能是歐洲社會第一次提到這個名字。這一次,這隻黑猩猩在航程中倖存下來,它與人類的相似度和模仿人類行為(例如用杯子喝茶)的能力給幾位英國學者留下了深刻印象。
瑞典博物學家卡尔·林奈於 1758 年出版了其著作《自然系統》之第十版,該書是生物分類學之基礎。他處理猿類的報告時將它們分為兩類:與人類更相似的報告被歸類為Homo troglodytes的另一個物種,而與猴子更相似的報告則被歸類為simia satyrus 的一個物種。物種名稱“troglodytes”起源於傳說中生活在非洲的野生人類,字面意思是“洞穴居民”,這個名字特別不合適,因為黑猩猩不住在洞穴裡。德國博物學家 約翰·弗里德里希·布盧門巴赫 認為林奈錯誤地將穴居人與之聯繫起來,並於 1776 年將其歸入 Simia troglodytes(現金已不再使用)。1816 年,德國博物學家 洛倫茲·奧肯 提出了一個折衷方案,他將穴居人歸為一個名為Pan的屬,以希臘神話中的潘神命名,這種分類至今仍被接受。
1863 年,托馬斯·亨利·赫胥黎發表論著《人類在自然中的地位的證據》(英語:Evidence as to Man's Place in Nature)。他將黑猩猩與其他人科非人類物種包括在一起—大猩猩、紅毛猩猩和長臂猿,並展示了它們的骨骼與人類骨骼的高度相似之處。赫胥黎和達爾文認為黑猩猩和大猩猩是最接近人類的兩個物種,因此建議在非洲尋找它們共同祖先的化石,因為當地是黑猩猩的起源地;此一預測在 20 世紀被調整。
20 世紀上半葉,幾位心理學家研究了被圈養的黑猩猩。這些研究主要記錄了它們的認知能力,並表明它們的智力超過了有尾猴。德國心理學家沃爾夫岡·苛勒於1913年至1917年在加那利群島的特內里費島建立了黑猩猩研究站,並出版了關於它們的書。他主要對它們藉由將多個組件組合在一起來解決問題的能力感興趣。例如,為了拿到一根用項圈掛在天花板上的香蕉,它們爬上幾個疊在一起的板條箱,然後用一根由兩根較短的棍子拼成的棍子敲打香蕉。苛勒相信他們藉由觀察和計劃得出了這些解決方案,而且不僅僅是反覆試驗,根據美國心理學家愛德華·桑代克的說法,這是動物能夠解決問題的唯一途徑。黑猩猩有時甚至會找到意想不到的解決方案,例如,當其中一隻黑猩猩用懇求的表情拉住苛勒的手臂時。苛勒很想跟著它,看看它要幹什麼,直到黑猩猩敏捷地爬過他的頭頂,抓住了香蕉,他才注意到自己正被直接領到香蕉下面。位於萊比錫的馬克斯·普朗克學會演化生物學研究所的靈長目動物研究中心現在以苛勒的名字命名。
1920年代,美國心理學家罗伯特·耶基斯在古巴建立了一個黑猩猩棲息地。他先是買了兩隻自家養的黑猩猩幼仔,睡在臥室裡,坐在餐桌旁,用餐具吃飯。1924年,他出版了一本關於它們的書 Almost Human。隨後幾年,耶基斯創立了耶魯大學的黑猩猩繁殖和研究中心,以及由洛克菲勒基金會資助的佛羅里達研究站,該研究站在他去世後被遷至埃默里大學,並仍以他的名字命名(參見耶基斯國家靈長類研究中心)。
然而,直到1960年代,由於難以進入熱帶雨林和難以觀察懼怕人類的野生猿猴,人們對野生黑猩猩的生活以及其他密切相關物種的生活幾乎一無所知。古人類學家路易斯·李奇了解這對研究人類演化的重要性,因此派遣三名女性研究人員研究三種人科非人類動物在自然環境中的生活—黛安·佛西到盧旺達研究大猩猩,碧露蒂·高蒂卡絲研究婆羅洲的猩猩,珍·古德研究坦桑尼亞的黑猩猩。珍·古德是李奇的秘書,且無大學學歷。當她 1960 年到達坦噶尼喀湖沿岸的貢貝河地區時,她在動物研究方面的經驗非常有限。在她執行任務的頭幾個月裡,她連找到黑猩猩都遇到了困難,它們經過很長時間才允許她近距離觀察它們。但就在李奇能夠從國家地理學會獲得的資助結束前不久,珍·古德有了兩個轟動性的發現:當地的黑猩猩會獵殺其他動物作為食物,它們會使用工具,甚至改造它們以適應自己的角色。在接下來的幾年裡,當地成為了一個國家公園,珍·古德將她的餘生奉獻給了研究當地黑猩猩的生活,並致力於推廣其公共關係以拯救和保護野生物種。她寫的幾本關於當地黑猩猩生活的書和製作的紀錄片在公眾和科學界都產生了巨大影響。
在中部非洲的 40 多個地點,目前正在對在野外發現的黑猩猩的生活進行實地研究,其中長期研究項目多年來一直追蹤同一黑猩猩群體:
- 坦桑尼亞的貢貝溪國家公園,珍·古德於 1960 年在此處開始了個人研究,並由她的學生繼續進行研究。
- 坦桑尼亞的馬哈萊山脈國家公園,是日本靈長目動物學家西田利貞等京都大學研究人員的研究項目所在地,有關研究始於1965年。
- 位於象牙海岸的塔伊國家公園,來自瑞士的研究員 Christophe 和 Hedwige Boesch 自 1976 年以來一直在這裡開展研究項目。自1990年代以來,他們一直隸屬於萊比錫的馬克斯·普朗克學會演化生物學研究所。
- 烏干達基巴萊國家公園生活著不少於 1,200 隻黑猩猩,其中四組已經習慣了研究人員和遊客的存在。該公園開展了兩個長期研究項目:自 1987 年以來在英國靈長目動物學家 Richard Wrangham(珍·古德的學生之一)的領導下在康亞瓦拉地區,以及自 1995 年以來在附近的 Ngogo 地區,由來自耶魯大學的美國人類學家 David Watts 領導。
- 塞內加爾的 Fongoli 地區,美國靈長目動物學家 Jill Pruetz 自 2001 年以來一直在這裡進行研究。根據古人類學領域的研究結果,該地區的獨特之處在於,它可以被用以研究疏林草原和森林棲息地中黑猩猩的生活—古人類據推斷也曾在類似條件下生活。與此同時,圈養、動物園和研究機構繼續研究黑猩猩的智力和社會行為,例如Royal Burgers' 動物園和耶基斯國家靈長類研究中心的荷蘭動物行為學家和靈長目動物學家弗蘭斯·德瓦爾的研究,萊比錫馬克斯·普朗克學會的美國心理學家 Michael Tomasello,以及京都大學的松澤哲郎和其他日本靈長目動物學家。
科學和醫學實驗
由於它們與人類的非常相似,黑猩猩是人類實驗最接近的替代品,因此多年來一直用於動物實驗,主要用於醫學目的,儘管與其他實驗動物相比數量相對較少,因為它很難將它們圈養起來且代價高昂。例如,黑猩猩在第一次太空飛行中先於人類宇航員(參看漢姆 (黑猩猩)),當時還不清楚失重和其他風險的影響是什麼。從1950年代開始,美國空軍為此從非洲進口了65個黑猩猩幼仔。其中,只有兩個最終在 1960 年代初期作為水星計劃的一部分被送進太空。它們的安全返回直接導致美國首位太空人艾倫·雪帕德的出現。其他黑猩猩被轉入研究等用途,其中包括參與了學習手語實驗的華秀。
多年來,公眾反對對黑猩猩進行實驗的呼聲越來越高,如今對黑猩猩進行合法醫學研究的僅有兩個國家—美國和加蓬。[27]1980 年代,美國用於實驗的黑猩猩數量在圈養繁殖成功後有所增加,這使得在不危及野外物種生存的情況下使用它們成為可能。但從那以後,其個體數又逐漸減少,尤其是在發現對它們的研究對尋找艾滋病疫苗和治療方法的貢獻微乎其微之後。反對實驗的Humane Society of the United States估計,1995 年美國參加實驗的黑猩猩數量約為 1800 個,2001 年為 1540 個,分佈在 13 個研究機構(根據美國國家衛生院的報告),並在 2007 年在九個研究機構中約 1,200個。[28]今天對黑猩猩的實驗,類似於對其他脊椎動物的實驗,大多在有關當局之動物福利機構的密切監督下。
在一些目前從事黑猩猩研究的科學機構中,例如耶基斯國家靈長類研究中心、萊比錫的馬克斯·普朗克學會和京都大學,黑猩猩永久性地群居在一起,有時在條件類似於自然條件的大型露天庭院中,它們自願參加各種實驗。研究人員將實驗設計得很有吸引力,藉由呼喊黑猩猩的名字邀請其參與,完成後甚至請求它們去邀請其他黑猩猩。習慣上,黑猩猩會因成功的表現而獲得食物獎勵,但京都大學的雌性個體“E”和雄性幼仔“Yumo”可能會在電腦螢幕前坐上數小時來解決記憶和數學挑戰,而沒有任何獎勵。
生存威脅與保護
黑猩猩被國際自然與自然資源保護聯盟定義為瀕危物種,僅次於“在野外滅絕但仍生活在圈養中”和“極度瀕危”。中非亞種受到的威脅最小—其種群數量估計為 80,000 個個體,生活在加蓬和剛果,但據估計正以每年 4.7% 的比例減少。東非亞種的數量估計為 13,000 個個體(但這是一個非常粗略的估計),而西非亞種的數量不超過 12,000 個個體,分散在一些偏遠地區。許多威脅共同嚴重破壞了該物種的野生種群,主要是人類捕獵、棲息地破壞和流行病。
中部非洲多個國家的法律禁止或限制獵殺黑猩猩,但其中許多國家的政局不穩意味著法律往往得不到執行。人們獵殺黑猩猩是為了獲取食物,也是為了將幼仔作為非法寵物出售。近幾十年來,小型武器的供應量不斷增加,導致非法狩獵的殺傷力增加。黑猩猩比其他靈長目動物更容易被獵殺,因為它們的體型較大,這使它們成為更大的目標,而且由於幼仔長期依賴母親,因此母親的死亡通常也會導致它們的死亡或被捕。
由於開墾土地用於農業、過度放牧、砍柴生火、工業發展和城市化,森林砍伐破壞了中部非洲黑猩猩的棲息地。林木的損失只是短期的破壞,風化、水土流失、物種多樣性的破壞、水源的消失等長期的破壞可能更為嚴重。2002年估計約有70%的黑猩猩生活區受到此類風險影響,假設中部非洲人口增長率保持不變,到2030年將有90%以上受到影響。
由於黑猩猩與人類的基因接近,它可能會感染許多危害人類並由人類傳播的傳染病,例如埃博拉、脊髓灰質炎、肺結核、水痘、肺炎、流感和感冒。這些疾病的流行影響了很大一部分現有的黑猩猩種群,並可能摧毀整個社群。1966 年,在珍·古德位在貢貝溪一帶的研究地區,一場小兒麻痺症疫情導致當地黑猩猩群體中很大一部分幼仔死亡,許多成年黑猩猩因此殘疾。與人的接觸越來越密切,其中不乏遊客來自其他地區,可能會增加感染此類疾病的風險。流行病也可能朝相反的方向傳播—根據一個共同的假設, HIV-1是導致人類艾滋病的更常見的HIV病毒形式,它是從一種起源於黑猩猩的相似但毒性較低的病毒進化而來的,並且很可能經由人類食用野生黑猩猩肉進入人體。[29]
對人類的襲擊行為
成年黑猩猩是強大而危險的野生動物,它們對人類的致命攻擊有記錄,儘管比鱷魚、獅子和河馬等非洲其他野生動物對人類的攻擊要少得多。在非洲的大部分棲息地,黑猩猩都非常懼怕人類並儘量避免靠近,但少數已經失去這種恐懼的黑猩猩可能主要攻擊人類幼童,它們像捕食猴子幼仔一樣捕殺這些人類幼童。在貢貝溪附近地區,珍·古德研究的群體中一隻特別大且好鬥的雄性綁架並捕食了一個人類嬰兒,這個嬰兒由該地區的一位年輕母親帶著,儘管禁止攜帶孩子進入當地。該雄性個體也曾襲擊珍·古德。
2009 年康涅狄格州發生了一起圈養黑猩猩的著名案例,當時一個被當作寵物飼養的成年雄性襲擊了飼主的友人,導致遇襲者身體嚴重變形且腦部受損,最後被警察開槍射殺。這隻黑猩猩似乎受到了阿普唑侖的影響,阿普唑侖是一種抗焦慮藥,對人類會產生幻覺的副作用。
在大眾文化中
許多黑猩猩現在還出現在馬戲表演、街頭表演、電影和電視節目等場合,主要是娛樂和各種脫口秀節目。它們幾乎總是扮演喜劇角色和小丑的原型角色,通常穿著人類服裝,藉由面部表情和模仿人類行為來取悅觀眾。這些節目的參與者幾乎都是幼仔,它們個頭小、友好、長相可愛。一個著名的例子是泰山電影中的角色,從 1930 年代開始由一系列黑猩猩演員扮演。動物權利組織與保育組織反對這種用途,因為黑猩猩通常來自非洲的非法貿易,那裡的母親被獵殺以獲取食物,幼仔被當作寵物出售。隨著這些黑猩猩成年,它們變得危險且難以控制,常在動物園結束他們的職業生涯。
科幻小說是少數黑猩猩在其中扮演重要角色的作品類型。最初例子可能是羅伯特·海萊因 1949 年的中篇小說,其中一隻黑猩猩的智力是藉由基因工程開發。具有人類智能的人科非人類動物的主題在皮埃爾·布勒 1964 年的小說《人猿星球》中得到發展,該小說曾多次被改編成電影。2001年版本中,海倫娜·寶漢·卡特飾演一隻聰明的黑猩猩,2011年版本中,劇情受到猿類語言實驗的影響,安迪·瑟克斯飾演基因工程黑猩猩“凱撒”一角。在1982年虛構小說《上帝的恩典》(英語:God's Grace)中,伯納德·馬拉默德描述了地球上最後一個人的命運,他領導著一個學習繁榮的黑猩猩群落,甚至與群裡唯一的黑猩猩建立了家庭,背景是人類的毀滅。1988年獲得雨果獎的大衛·布林的小說The Uplift War中,主人公之一是基因工程黑猩猩Fiben Bolger,它的名字取自珍·古德著作中的一個黑猩猩。
人類演化與黑猩猩的關係
參考文獻
- Humle, T., Maisels, F., Oates, J.F., Plumptre, A. & Williamson, E.A. . The IUCN Red List of Threatened Species 2016. [2018-07-27].
- . [2016-08-17]. (原始内容存档于2016-08-19).
- . IUCN Red List of Threatened Species. 國際自然保護聯盟. [2023-03-12]. (原始内容存档于2023-06-07) (英语).
- Walker, A. (2009) "The strength of great apes and the speed of humans" Current Anthropology .
- McBrearty, Sally; Jablonski, Nina G. . Nature. 2005-09, 437 (7055). ISSN 1476-4687. doi:10.1038/nature04008 (英语).
- Waterson, Robert H.; Lander, Eric S.; Wilson, Richard K.; The Chimpanzee Sequencing and Analysis Consortium. . Nature. 2005-09, 437 (7055) [2023-03-12]. ISSN 1476-4687. doi:10.1038/nature04072. (原始内容存档于2021-05-25) (英语).
- Hey, Jody. . Molecular Biology and Evolution. 2010-04, 27 (4) [2023-03-12]. ISSN 1537-1719. PMC 2877540
 . PMID 19955478. doi:10.1093/molbev/msp298. (原始内容存档于2023-03-12).
. PMID 19955478. doi:10.1093/molbev/msp298. (原始内容存档于2023-03-12). - Gonder, Mary Katherine; Locatelli, Sabrina; Ghobrial, Lora; Mitchell, Matthew W.; Kujawski, Joseph T.; Lankester, Felix J.; Stewart, Caro-Beth; Tishkoff, Sarah A. . Proceedings of the National Academy of Sciences. 2011-03-22, 108 (12) [2023-03-12]. ISSN 0027-8424. PMC 3064329 . PMID 21368170. doi:10.1073/pnas.1015422108. (原始内容存档于2023-03-12) (英语).
- Wilson, M. . www.semanticscholar.org. 2013 [2023-03-12]. (原始内容存档于2023-03-12) (英语).
- Henke, Winfried; Tattersall, Ian (编). . (PDF). Springer Berlin, Heidelberg. 2015 [2023-03-12]. ISBN 978-3-642-39978-7. doi:10.1007/978-3-642-39979-4. (原始内容存档 (PDF)于2016-08-17) (英语).
- Crockford, Catherine; Boesch, Christophe. . Behaviour. 2005, 142 (4) [2023-03-12]. ISSN 0005-7959. (原始内容存档于2023-03-12).
- Wroblewski, Emily E.; Murray, Carson M.; Keele, Brandon F.; Schumacher-Stankey, Joann C.; Hahn, Beatrice H.; Pusey, Anne E. . Animal Behaviour. 2009, 77 (4) [2023-03-12]. ISSN 0003-3472. PMC 2689943 . PMID 19498952. doi:10.1016/j.anbehav.2008.12.014. (原始内容存档于2023-03-12).
- McGrew, W. C. (2010). Chimpanzee technology. Science, 328(5978), 579–580. https://doi.org/10.1126/science.1187921
- Haslam, Michael; Hernandez-Aguilar, Adriana; Ling, Victoria; Carvalho, Susana; de la Torre, Ignacio; DeStefano, April; Du, Andrew; Hardy, Bruce; Harris, Jack; Marchant, Linda; Matsuzawa, Tetsuro. . Nature. 2009-07, 460 (7253) [2023-03-12]. ISSN 1476-4687. doi:10.1038/nature08188. (原始内容存档于2023-09-13) (英语).
- Boesch, Christophe; Head, Josephine; Robbins, Martha M. . Journal of Human Evolution. 2009-06, 56 (6) [2023-03-12]. ISSN 1095-8606. PMID 19457542. doi:10.1016/j.jhevol.2009.04.001. (原始内容存档于2023-03-28).
- Pruetz, Jill D.; Bertolani, Paco. . Current Biology. 2007-03-06, 17 (5) [2023-03-12]. ISSN 0960-9822. doi:10.1016/j.cub.2006.12.042. (原始内容存档于2023-06-29) (英语).
- Pruetz, J. D.; Bertolani, P.; Ontl, K. Boyer; Lindshield, S.; Shelley, M.; Wessling, E. G. . Royal Society Open Science. 2015-04, 2 (4) [2023-03-12]. ISSN 2054-5703. PMC 4448863 . PMID 26064638. doi:10.1098/rsos.140507. (原始内容存档于2023-01-10) (英语).
- Horner, Victoria; de Waal, Frans B. M. . Progress in Brain Research. 2009, 178 [2023-03-12]. ISSN 1875-7855. PMID 19874958. doi:10.1016/S0079-6123(09)17801-9. (原始内容存档于2023-03-12).
- Luncz, Lydia V.; Mundry, Roger; Boesch, Christophe. . Current Biology. 2012-05-22, 22 (10) [2023-03-12]. ISSN 0960-9822. doi:10.1016/j.cub.2012.03.031. (原始内容存档于2023-03-12) (英语).
- Biro, D.; Matsuzawa, T. . Animal Cognition. 2001-11, 4 (3-4) [2023-03-12]. ISSN 1435-9448. PMID 24777509. doi:10.1007/s100710100086. (原始内容存档于2023-03-12).
- Martin CF, Bhui R, Bossaerts P, Matsuzawa T, Camerer C. Chimpanzee choice rates in competitive games match equilibrium game theory predictions. Sci Rep. 2014 Jun 5;4:5182. doi: 10.1038/srep05182. PMID 24901997; PMCID: PMC4046491.
- Proctor, Darby; Williamson, Rebecca A.; de Waal, Frans B. M.; Brosnan, Sarah F. . Proceedings of the National Academy of Sciences. 2013-02-05, 110 (6) [2023-03-12]. ISSN 0027-8424. PMC 3568338 . PMID 23319633. doi:10.1073/pnas.1220806110. (原始内容存档于2023-03-12) (英语).
- Gallup, Gordon G. . Science. 1970-01-02, 167 (3914) [2023-03-12]. ISSN 0036-8075. doi:10.1126/science.167.3914.86. (原始内容存档于2023-10-25) (英语).
- Call, Josep; Tomasello, Michael. . Trends in Cognitive Sciences. 2008-05, 12 (5) [2023-03-12]. ISSN 1364-6613. PMID 18424224. doi:10.1016/j.tics.2008.02.010. (原始内容存档于2023-03-12).
- Schmelz, Martin; Call, Josep; Tomasello, Michael. . Proceedings of the National Academy of Sciences. 2011-02-15, 108 (7) [2023-03-12]. ISSN 0027-8424. PMC 3041140 . PMID 21282649. doi:10.1073/pnas.1000469108. (原始内容存档于2023-05-10) (英语).
- . 明報. 2014-07-14 [2014-07-15]. (原始内容存档于2014-07-17).
- Waal, Frans B. M. de. . PLOS Biology. 2012-03-27, 10 (3) [2023-03-12]. ISSN 1545-7885. PMC 3313912 . PMID 22479150. doi:10.1371/journal.pbio.1001291. (原始内容存档于2023-03-12) (英语).
- Conlee, K. . www.semanticscholar.org. 2008 [2023-03-12]. (原始内容存档于2023-03-12) (英语).
- Keele, Brandon F.; Van Heuverswyn, Fran; Li, Yingying; Bailes, Elizabeth; Takehisa, Jun; Santiago, Mario L.; Bibollet-Ruche, Frederic; Chen, Yalu; Wain, Louise V.; Liegeois, Florian; Loul, Severin. . Science (New York, N.Y.). 2006-07-28, 313 (5786) [2023-03-12]. ISSN 1095-9203. PMC 2442710 . PMID 16728595. doi:10.1126/science.1126531. (原始内容存档于2023-03-24).