原賴氏龍屬
原賴氏龍(屬名:,意為「破曉的賴氏龍亞科」)是植食性鴨嘴龍超科的一屬恐龍,來自白堊紀晚期之初的美國。底下包含單一物種:卡蘿瓊斯原賴氏龍(E. caroljonesa),於1998年由古生物學家詹姆斯·科克蘭所命名。原賴氏龍的正模標本是在1993年由卡蘿和拉瑪·瓊斯(Carole and Ramal Jones)發現,種名即紀念前者。自此之後,包括成體與幼體的數百具骨骼相繼被發現,幾乎涵蓋整副骨架的所有部位。所有標本都是來自猶他州艾麥里縣的雪松山組穆森圖奇段(Mussentuchit Member)地層。
| 原赖氏龙属 | |
|---|---|
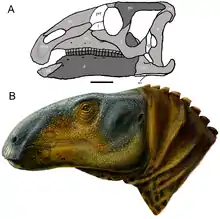 | |
| 頭骨重建圖(A)與頭部復原圖(B) | |
| 科学分类 | |
| 界: | 动物界 Animalia |
| 门: | 脊索动物门 Chordata |
| 纲: | 蜥形纲 Sauropsida |
| 总目: | 恐龍總目 Dinosauria |
| 目: | †鸟臀目 Ornithischia |
| 亚目: | †鸟脚亚目 Ornithopoda |
| 演化支: | †禽龍類 Iguanodontia |
| 演化支: | †橡树龙形态类 Dryomorpha |
| 演化支: | †直拇指龍類 Ankylopollexia |
| 演化支: | †硬棘龍類 Styracosterna |
| 演化支: | †鴨嘴龍形類 Hadrosauriformes |
| 总科: | †鴨嘴龍超科 Hadrosauroidea |
| 属: | †原赖氏龙属 Eolambia Kirkland, 1998 |
| 模式種 | |
| †卡蘿瓊斯原賴氏龍 Eolambia caroljonesa Kirkland, 1998 | |
原賴氏龍是家族中的大型物種,身長約6公尺。雖然牠與亞洲的鴨嘴龍超科如馬鬃龍、原巴克龍、喬伊爾齒龍近緣,其頭骨、脊椎、肢骨特徵顯示,可能其實與北美洲的始鴨嘴龍更為近緣。根據方骨和肩胛骨的筆直度判斷這群物種構成鴨嘴龍超科一群單系、特有種的輻射適應演化支。雖然原賴氏龍與鴨嘴龍科、尤其是賴氏龍亞科有諸多相似特徵,導致最初曾被鑑定為其中一員,但後來這些相似性都被否定,可能只是趨同演化或錯誤鑑定。
原賴氏龍可能生活在潮濕氾濫平原的湖泊邊緣森林環境,以蕨類、裸子植物、開花植物為食。從穆森圖奇段沉積物中的交替帶反映出,湖泊水平面隨時間演進而變化,是因為地球歲差引起的周期性乾濕循環。原賴氏龍在幼年時可能被居住在湖泊水域的大型鱷形超目所獵捕。隨著年齡增長,體型將使鱷類無法再構成威脅,而約八至九歲的成熟個體將被如新獵龍科的西雅茨龍之類的大型獸腳類所掠食。
發現歷史
最初的發現與敘述

1979年彼得·高爾頓和詹姆斯·詹森敘述了一塊破碎的右股骨,標本編號BYU 2000,屬於一種鴨嘴龍類恐龍,發現於猶他州拱門國家公園中屬於雪松山組的沉積層中。雖然保存很差,卻與同一論文敘述的第二個北美股骨一起,成為全世界第一個下白堊紀鴨嘴龍類標本,意義重大。高爾頓和詹森推測未來將能在該地層發現更完整的鴨嘴龍類遺骸。[1]在猶他州艾麥里縣代爾堡附近的聖拉斐爾湧浪西部地區的小型脊椎動物採石場也發現了數種鴨嘴龍類牙齒,並於1991年由傑麥可·帕里許(J. Michael Parrish)敘述。後來在1993年,卡蘿·瓊斯(Carole Jones)和她的丈夫拉瑪·瓊斯(Ramal Jones)在湧浪西北地區的化石點發現了破碎骨骼。他們將該地點引起了唐諾·伯格(Donald Burge)的注意,他是當時東猶他歷史博物館大學(CEUM)的校長。這個過去被稱為「CEUM Locality 42em366v」的化石點,於是就被改名為「卡蘿點(Carol's Site)」已作紀念。化石目前保存為編號CEUM 9758標本,是一隻成年鴨嘴龍類的部分骨骸,包括部分頭骨、脊椎、坐骨、腿部。CEUM 5212發現於卡蘿點附近,是一個成體的部分頭骨與前肢。[2]CEUM 8786後來也在卡蘿點發現,是一個成體左股骨,直到2012年之前都未被敘述。[3]
與此同時,奧克拉荷馬自然史博物館(OMNH)在湧浪西南處的挖掘活動發現了屬於同種鴨嘴龍類的遺骸。[2]OMNH發現的標本起初包括來自五個地點的六隻個體:OMNH v237地點有兩個幼體骨骼,包含脊椎、肩胛骨、尺骨、髂骨、坐骨以及後肢材料;OMNH v824地點有一個部分幼體骨骼,包含部分頭骨、前後肢與一個背椎;OMNH 27749標本是來自OMNH v696地點的薦骨和坐骨;OMNH 24389標本是來自OMNH v214地點的坐骨;OMNH 32812標本包含來自OMNH v866地點的肩胛骨、兩個尾椎及其他尚未挖出的材料。OMNH也在數個採石場進一步發現了鴨嘴龍類牙齒。OMNH的理察·西費利(Richard Cifelli)授權讓詹姆斯·科克蘭來研究採集到的標本。[2]科克蘭於1994年古脊椎動物協會的年度研討會會議摘要中,對標本進行了初步敘述。[4]

1998年,科克蘭在新墨西哥自然歷史與科學博物館公告發表的論文中正式將標本命名為新屬原賴氏龍及其模式種兼唯一種卡蘿瓊斯原賴氏龍(Eolambia caroljonesa)。屬名由希臘文字首eos/ἠώς(「黎明、破曉」,代表「早期」之意)加上後綴lambia(源自於賴氏龍,以加拿大古生物學家勞倫斯·賴博為名)構成。整個屬名的意思是「早期的賴氏龍亞科」,代表可能屬於最基礎的賴氏龍亞科位置。而種名紀念發現者卡蘿·瓊斯。原賴氏龍的名字是由古生物藝術家麥克·史奎普尼克所提名,以取代科克蘭在1998年論文之前原先使用的非正式名稱卡蘿瓊斯原鴨嘴龍("Eohadrosaurus caroljonesi")。[2]
後續發現

自1998年起陸續有許多挖掘活動發現更多屬於原賴氏龍的骨骼。牠現在成為森諾曼階至桑托階時期所知最完整的禽龍類。已採集的化石幾乎涵蓋整個骨架的每個部位。在艾麥里縣南邊的採石場發現了至少十二隻個體,包括至少八隻幼體,有頭骨及其他身體骨骼。[5]該採石場被稱為「西費利2號卡蘿瓊斯原賴氏龍採石場(Cifelli #2 Eolambia caroljonesa Quarry)」,或較正式的稱作「CEUM locality 42em432v」。艾麥里縣南邊另一處採石場「柳泉8號採石場(Willow Springs 8 quarry)」/「CEUM locality 42Em576v」保存了至少四隻幼體。來自這些採石場的化石於2012年由科克蘭與安德魯·麥當勞(Andrew McDonald)、約翰·伯德(John Bird)、彼得·多德森一同敘述,同時根據新標本列出了更新版的鑑定特徵。[3]
西費力2號採石場發現的一具標本,編號CEUM 34447的右齒骨,與其他原賴氏龍成體及幼體標本相比之下,顯得不太尋常。在其他個體中,齒骨中心點深度與總長度比值通常介於0.19至0.24,CEUM 34447卻有0.31異常地深。齒骨前端也沒有明顯變深,於是缺乏原賴氏龍齒骨的其中一項鑑定特徵。然而標本有另一個鑑定特徵,為下頜骨喙狀突(coronoid process)前端擴張。雖然這個標本可能是原賴氏龍的一隻變異個體,麥當勞等人表示牠可能是另一種與原賴氏龍齒骨形態不同的獨特鴨嘴龍類(但採石場中的任何其他材料都沒有證據支持)。於是他們將此標本視為鴨嘴龍類未定物種。[3]

2009年至2010年期間,在猶他地質調查局的同意下,菲爾德自然史博物館也在艾麥里縣南方展開了考察活動。這次挖掘在「FMNH locality UT080821-1」/「Akiko's Site」(由Akiko Shinya於2008年發現的化石點)進行。過程中挖出了167塊關節脫落、彼此關聯靠近的骨骼,並包裹在兩塊石膏中採集下來。其中一塊石膏包括骨盆與脊椎,標本編號為FMNH PR 3847;於2017年由麥當勞與泰瑞·蓋茨(Terry Gates)、林塞·贊諾(Lindsay Zanno)、彼得·馬克維奇(Peter Makovicky)一同敘述。另一個由菲爾德博物館發現的化石點為「三峰採石場(Triple Peak quarry)」/「FMNH locality UT130904-2」,產出超過400塊骨骼、至少四隻個體的骨床。[6]
地質定年

所有原賴氏龍的發現地都屬於雪松山組的穆森圖奇段(Mussentuchit Member),其成分主要由交錯的淺灰色至灰色泥質砂岩及泥岩層構成,同時也顯示出兩個獨立河道系統的沉積物。系統下方由四層構成,分別為:泥質砂岩、細粒砂岩、粉沙質泥岩、細粒砂岩,後兩層為植物殘留物。西費力2號採石場與此系統有所關聯。上部系統為綠色細粒砂岩。穆森圖奇段還有三層屬於火山灰沉積物的皂土黏土層,分別為白灰色、黃色、棕褐色。這幾層中間部分穿過西費力2號採石場。[5]皂土亦出現於整個穆森圖奇段砂岩中。[6]西費力2號採石場的骨骼本身被染黑,由方解石、石英、黃鐵礦、碳酸根-羥磷灰石所取代。[5]
孢粉學[7][8][9]、巨植物群及無脊椎動物研究[10]起初認為雪松山組最上層(含穆森圖奇段)年代可追溯至阿爾比階。[2]1997年由西費力等人所作的雪松山組上層火山灰中的透長石結晶氬-氬定年得出年代為9700萬±100萬年前,當時被認為在阿爾比階至森諾曼階交界不久之後,與上覆的達柯塔組為早森諾曼階相近。[11]後來於2007年,詹姆斯·蓋瑞森(James Garrison)等人將通過西費力2號採石場的火山灰層(可能與先前西費力等人鑑定的為同一層)定年為9670萬±500萬年前。在大致接近的水平面上,兩項研究估計彼此一致,支持了原賴氏龍來自森諾曼階。蓋瑞森等人還重新分析整個穆森圖奇段的花粉結構,得出整體年代橫跨了阿爾比階-森諾曼階交界。[5]
敘述

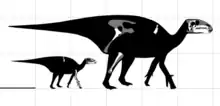
原賴氏龍是種大型鴨嘴龍超科。最初估計頭骨長度落在1公尺,[2]但這是被比例過長的鼻部所誤導,後來發現額外材料便被糾正。[3]2016年·保羅估計原賴氏龍身長6公尺及體重1噸,[12]與2012年·霍茨所估計的6.1公尺身長相符。[13]較早的2008年,一個成年個體標本被測量身長5.2公尺及臀高2公尺。[5]


頭骨
原賴氏龍不具頭冠的頭骨整體形狀與馬鬃龍、原巴克龍相似。吻部前端非常粗糙,由許多孔洞所穿透。每個前上頜骨尖端有兩個齒狀結構稱為小齒(denticles),也可見於近緣的原巴克龍。再往後前上頜骨下枝後部突然上突關住後面的鼻孔,如同原巴克龍、始鴨嘴龍及其他鴨嘴龍類。此部分關節與上頜骨兩根指狀突相連,類似始鴨嘴龍。上頜骨體本身不具凹陷或任何眶前孔的痕跡,如同馬鬃龍、始鴨嘴龍和其他鴨嘴龍類。其中一項用來鑑定原賴氏龍的特徵是上頜骨齒列側視輪廓呈凹形,近似於馬鬃龍、原巴克龍、數種其他鴨嘴龍形類,但與始鴨嘴龍不同。[3]


如同原巴克龍和其他鴨嘴龍類,上頜骨背側以指狀突與顴骨(構成眼眶及顳顬孔下緣)的凹槽吻合相連。顴骨下緣在顳顬孔水平以下有個堅硬凸緣,也可在馬鬃龍、原巴克龍、始鴨嘴龍、其他數種鴨嘴龍類觀察到。顴骨上方與眶後骨相連,其與眼窩相鄰處表面粗糙(如同始鴨嘴龍),但側面則顯得光滑。頭骨後部,方骨與鱗狀骨以一個上視呈D字形的關節相連。左右鱗狀骨可能彼此完全接觸,只有背側被頂骨的一個小突起分隔開來。構成頭骨後部上區的上枕骨平坦且近乎垂直,如同原巴克龍和其他鴨嘴龍類的情況。[3]

和前上頜骨一樣,前齒骨也有小齒。有個突出的內背側突起,是為片狀結構,也可於原巴克龍和其他鴨嘴龍類觀察到。數個額外的片狀小齒出現於內背突兩側,亦見於原巴克龍。前齒骨後面與齒骨(構成下頜主要部分)關節相連。在前齒骨與齒骨關節相連處之間、齒骨第一顆牙齒的位置有個小型牙間隙,也在馬鬃龍、原巴克龍和其他鴨嘴龍類觀察到。齒骨前部有特徵性的加深,如同始鴨嘴龍、豪勇龍、巴克龍。齒骨外表面有兩個突起,其中一個屬於喙突,如同原巴克龍和其他鴨嘴龍類。與始鴨嘴龍和其他數種鴨嘴龍類相似,但與原巴克龍不同的是,喙突只有前端加厚。如同馬鬃龍、原巴克龍和其他數種禽龍類,上隅骨側邊、位於齒骨後方處有個小孔。[3]
與其他禽龍類相似的是,原賴氏龍的牙齒排列呈緊密間隔與互相緊鎖的環列。無論何時,32個上頜骨齒槽每個都包含三顆牙齒,而30個齒骨齒槽每個都包含四顆牙齒。其中每個齒槽中的兩顆牙齒都是像原巴克龍一樣的汰換性牙齒,其他則是主要功能性的牙齒。每個功能性牙齒都有個磨耗面。就像原巴克龍、公婆泉龍、始鴨嘴龍和其他鴨嘴龍類,每個上頜骨齒冠只有一道嵴,略微偏向嘴部中線;每個齒骨齒冠的特徵為有個主嵴和一道靠近嘴部中線的副嵴,這種狀況也存在於始鴨嘴龍和其他鴨嘴龍類。所有齒冠的前後緣都長著細小的小齒狀鋸齒邊緣,也見於原巴克龍。[3]
脊柱


如同馬鬃龍、原巴克龍、公婆泉龍、其他硬棘龍類,原賴氏龍的頸椎大幅雙後凹(opisthocoelous),即前端大幅突出、後端大幅凹入。不尋常的是,第三頸椎前端位置稍高於後端,其他脊椎都未觀察到此情況。其他更後面的頸椎在加長的關節突之間有著更大角度,稱為後關節突(postzygapophyses)。橫突(transverse processes)也加長,上還有進一步的關節突:圓潤的附突(parapophyses)和桿狀上橫突(diapophyses)。在最後面的頸椎中,有個深凹將後關節突隔開,神經棘前緣陡峭,構成突出的尖刺狀突起。[3][6]
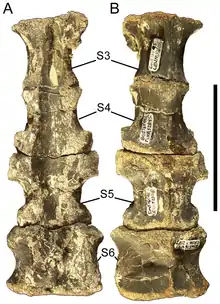
最前面幾節脊椎與頸椎相似,但神經棘較高、更像叉狀。其餘背椎為兩端平坦(amphiplatyan),除了神經棘較高、更偏矩形,後關節突的加長程度較少、更偏基座狀。附突為凹陷而非突起,由橫突位移至神經弓,位於橫突和關節突之間,稱為脊椎前部的前關節突(prezygapophyses)。然而在最後幾節背椎中,附突移回橫突基部。[3]在第三或第四背椎,附突位置非常接近神經即與椎體之間的縫線,與其他背椎不同,但和埃德蒙頓龍的前幾節背椎相似。分隔前關節突的中脊在後部背椎變得更加突出,也很像埃德蒙頓龍。[6]

在目前已知的原賴氏龍標本中,保存最佳的薦骨有七節脊椎。有鑑於這隻個體尚未成熟,而鴨嘴龍類薦椎數目會隨著年齡增長而增加,因此推測成體會有更多節薦椎。[2]尾椎為前後端都凹入的雙內凹(amphicoelous)形式。具有基座狀的前關節突與片狀的後關節突,後者由一道凹陷所隔開。在中部和後部尾椎,前關節突延長呈桿狀,而後關節突收縮。此外,神經棘從矩形轉變成大幅彎曲(前凹後凸),椎體更為加長,橫突也消失。[3][6]
肢骨和肢帶


如同原巴克龍,原賴氏龍的肩胛骨幾乎呈矩形,側邊直,與末端幾乎成直角。肱骨朝中線彎曲,肱骨上的三角肌嵴明顯增厚。肱骨與橈骨連接的關節外表面有個圓形隆起。尺骨與橈骨都相對筆直。尺骨頂部有三個刺,稱為鷹嘴突及兩個額外的三角形凸緣。同時橈骨頂端有個平坦的內表面,底端內表面有明顯的平坦處,兩處都與尺骨關節相連。手部有五指。掌骨中,第二掌骨彎曲,第三掌骨筆直,第四掌骨大幅彎曲。如同禽龍、原巴克龍以及其他基礎禽龍類,第一指爪呈現獨特的圓錐狀。第二及第三指爪呈蹄狀,其中第三指爪兩側有粗糙的凸緣,也是與其他禽龍類共有的特徵。[3]
髂骨預臼(preacetabular)(髖臼前面的位置)突具有水平向的靴狀,與突起底部構成一鈍角;此外,骨頂緣外突以構成骨底緣上位於髂骨-坐骨關節的環;此特徵亦見於原巴克龍和其他禽龍類。[3]以禽龍類的髂骨來說,髖臼本身相對大型。[14]延髂骨頂面出現一個獨特的凸緣,這個特徵與來自木藤組的未命名鴨嘴龍類共有。髂骨後臼(postacetabular)(位於髖臼之後)突後部漸細,頂緣無斷裂。恥骨前突向前擴展,如同原巴克龍、公婆泉龍和其他禽龍類。成體坐骨軸是典型地直(而幼體坐骨軸彎曲,為禽龍類較常見的狀況),軸底部有個收縮地恥靴向前擴展。[3][6]

股骨和肱骨一樣彎曲,但前者朝外。但股骨下半部典型地直。股骨中間的第四轉子縱向高且呈三角形,類似原巴克龍、公婆泉龍和其他禽龍類,但與更基礎的鳥腳類不同。脛骨腓骨和尺骨橈骨一樣也有直的軸,彼此關節互相連接;腓骨底部粗糙突起剛好符合脛骨底端脛嵴後兩個球狀髁之間的空間。脛骨頂部粗糙,代表股骨和脛骨之間有軟骨罩的存在。腳部有四趾。如同掌骨的情況,第二蹠骨彎曲,第三蹠骨直,第四蹠骨大幅彎曲。腳趾非常厚實,接近每根腳趾末端的最後一節趾骨都特別粗壯。如同手部的第二、三指,腳部第二、三、四趾寬闊平坦並呈蹄狀。[3]
分類
最初鑑定為鴨嘴龍科
在科克蘭最初的敘述中,他認為原賴氏龍屬於鴨嘴龍科,以大衛·魏尚沛、大衛·諾曼、丹·奎戈瑞斯庫(Dan Grigorescu)1993年的定義為依據。魏尚沛等人以七個共同特徵來定義鴨嘴龍科:①上頜骨升突向上擴張②缺乏將方骨與方軛骨分隔開來的副方孔(paraquadrate foramen)③隅骨在下頜內面的位置④上隅骨缺乏上隅孔⑤上頜骨牙齒狹窄⑥每個齒位長著三顆或更多顆牙齒⑦肩胛骨上緣的縮減。[15]在原賴氏龍中可觀察到第①⑤⑥⑦個特徵,其餘因材料缺乏而未知。科克蘭進一步將原賴氏龍歸入真鴨嘴龍類(Euhadrosauria),根據魏尚沛等人的定義,包含鴨嘴龍科內主要兩個分支:鴨嘴龍亞科和賴氏龍亞科的共同祖先、及其所有後代。[15]他的分類以五項特徵為依據,原賴氏龍全部都有:①前上頜骨有小齒突②方骨與下頜的關節狹窄③齒骨牙齒狹窄④每顆牙齒上都有單一邊緣隆突⑤肱骨三角肌嵴呈角度。[2]
在鴨嘴龍科中,科克蘭進一步認為原賴氏龍要麼是賴氏龍亞科的基礎物種、要麼與賴氏龍亞科互成姊妹群。他鑑定出原賴氏龍與賴氏龍亞科共有的六項特徵,為後來其他許多研究所沿用:[15][16][17][18]①前上頜骨缺乏孔洞②至少一部分鼻孔被前上頜骨包圍著③上頜骨板狀物的發育④尾椎神經棘非常高⑤肱骨的堅韌性⑥成體坐骨有大型靴狀底座。科克蘭發現另外兩項特徵將無頭冠的原賴氏龍與其他有頭冠的賴氏龍亞科區別開來,這些被認為與頭冠發育有關:①鼻腔高於眼眶②頂骨縮短。若原賴氏龍屬於賴氏龍亞科,則後者將需要被重新定義以排除以上兩項特徵。或者如果原賴氏龍是賴氏龍亞科的姊妹群,牠將代表著鴨嘴龍科中鴨嘴龍亞科與賴氏龍亞科之間的差異形態。這種可能性由原賴氏龍薦骨底部有溝槽所支持,科克蘭認為這是鴨嘴龍亞科的一項關鍵性特徵[17][18]——雖然甲龍類和角龍類也有這種趨同演化出的特徵。[2]
為了支持賴氏龍亞科與原賴氏龍的關聯性,科克蘭進行了一項種系發生學分析,其得出的演化樹將原賴氏龍對應放到賴氏龍亞科的基礎位置,如下所示。雖然科克蘭引用了證據作為支持,他還是指出了一項看起來不連貫的特徵。如同在他最初敘述的幼體髂骨上觀察到的情況,髂骨上用來限制股骨大轉子的突起物——反轉子(antitrochanter),比基礎鴨嘴龍科預期的發育程度還要更差。麥可·布雷特·瑟曼(Michael Brett-Surman)指出成體可能有發育較佳的反轉子。另一方面,進階型賴氏龍亞科亞冠龍就算在剛孵化階段即有發育良好的反轉子。科克蘭提出基礎鴨嘴龍科在生長過程中,反轉子的發育可能比進階鴨嘴龍科(如亞冠龍)來得更晚的可能性。[19]於是他表示原賴氏龍的成體標本將可解決此疑問。[2]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
重新鑑定為非鴨嘴龍科
2001年,傑森·黑德(Jason Head)根據奧克拉荷馬自然史博館保存的原賴氏龍標本重新檢驗其種系發生學位置。他意識到科克蘭提到原賴氏龍擁有所謂的賴氏龍亞科特徵不是錯誤就是可疑的。如黑德所述,前上頜骨其實有一個孔洞;前上頜骨僅有底緣與鼻孔接壤;上頜骨前面除了板狀物還有兩個指狀突起;尾椎的高神經棘也可在非賴氏龍亞科的豪勇龍和巴克龍身上觀察到;且坐骨末端擴張也可見於彎龍、禽龍、豪勇龍、計氏龍、巴克龍,代表這是禽龍類的祖徵,而到鴨嘴龍亞科才失去。[18]雖然ML·卡薩諾瓦斯(M.L. Casanovas)等人於1999年的研究將賴氏龍亞科描述為坐骨末端有擴張的靴狀突而非棒狀,[20]黑德反駁道冠龍和賴氏龍[21]的坐骨末端沒有像曼特爾龍或豪勇龍來得那麼擴張。[14][22]

黑德還證實原賴氏龍牙齒比科克蘭原先假設的更寬及更不對稱,並指出雖然科克蘭最初所述成體標本每個齒位有三顆或以上的牙齒是正確的[3],但亞成體每個齒位卻僅有兩顆牙齒,這與鴨嘴龍科不同。[14]黑德進一步表示原賴氏龍有七節薦椎,不像進階型鴨嘴龍科有八節或更多,且其反轉子發育程度比豪勇龍、高吻龍還要更差。[23]對OMNH標本的檢驗進一步顯示,髂骨後髖臼區域未構成明顯的突起,底部與股骨間的關節也不突出,此兩項特徵都將原賴氏龍與更進階的禽龍類如鴨嘴龍科區別開來。於是黑德得出結論為,原賴氏龍是鴨嘴龍超科的基礎物種,位於鴨嘴龍科之外,種系發生學分析並支持牠是與原巴克龍近緣的非鴨嘴龍科鴨嘴龍超科。[14]
後續研究

2004年,諾曼首次將原賴氏龍包含進一份全面性的禽龍類種系發生學分析,收錄於其著作《恐龙 (书籍)》第二版中關於禽龍類的章節。他發現原賴氏龍是高吻龍的姊妹群,兩者都比始鴨嘴龍、原巴克龍、鴨嘴龍科還基礎。[24]同一章節中,傑克·霍納、魏尚沛、凱瑟琳·佛斯特另外對鴨嘴龍科進行種系發生學分析,同樣包含進原賴氏龍。他們再次發現原賴氏龍的確是非鴨嘴龍科的鴨嘴龍超科,但這次將牠與始鴨嘴龍互成姊妹群。雖然黑德根據共有衍徵定義特別重新定義了鴨嘴龍科來包含進始鴨嘴龍,[14][25]霍納和同事則採用了另一項以分類學為準則的定義,將始鴨嘴龍與原賴氏龍一併排除在外。他們還定義了將原賴氏龍與其他鴨嘴龍科區別開來的特徵:齒骨牙齒上有粗糙的小齒突、喙突略微擴張。[26]
在接下來的幾年中,原賴氏龍的種系發生學分析持續存在著差異。在2009年漢斯·戴爾特·蘇伊士和亞歷山德·阿維葉諾夫(Alexander Averianov)敘述列弗尼斯氏龍的論文中,發現相較於高吻龍和原巴克龍,始鴨嘴龍佔據著中間程度的位置,並與福井龍互成姊妹群。[27]與此同時,2010年亞伯特·皮耶妥·馬奎茲(Albert Prieto-Márquez)和馬克·諾瑞爾以霍納等人的結論作為基礎的研究發現原賴氏龍反而才是始鴨嘴龍的姊妹群。他們還指出原賴氏龍有哪些特徵是與鴨嘴龍科趨同演化的:牙齒上單一隆突的存在;前齒骨外角大致接近方形;與方軛骨關節相連的溝槽中心點大致位在方骨表面的一半位置(由上而下低於骨頭高度60%)。[28]2012年,霍茨將原賴氏龍與高吻龍一同歸類為直拇指龍類的原始物種。[13]

2012年,麥當勞對禽龍類進行了種系發生學分析,當中參考了原賴氏龍的新標本資訊。與過去研究不同的是,他發現原賴氏龍與原巴克龍互成姊妹群。這樣的近緣關係是以兩項共有衍徵為依據:方骨除了後彎的頂端外,整體上是呈筆直的;肩胛骨頂緣和底緣近乎平行。[3]2014年鄭文傑等人在敘述薄氏龍新標本的研究中,將金塔龍也納入以上姊妹群。[29]而2015年何塞·珈斯卡(José Gasca)在德拉帕倫特龍的種系發生學研究中,則採納麥當勞原本的分類版本。[30]但之後麥當勞進一步大幅修改了他的種系發生學資料庫,為準備於2017年描述新的FMNH標本。如同霍納等人、皮耶妥·馬奎茲與諾瑞爾的研究,麥當勞等人發現原賴氏龍與始鴨嘴龍互成姊妹群,兩者都位於鴨嘴龍形態類的基礎位置,結果如下方支序圖所示。[6]依照麥當勞2012年版的資料庫,法蘭西斯科·韋篤(Francisco Verdú)於2015年及2017年的禽龍分類研究中獨立完成了相似的結果:[31][32]原賴氏龍與始鴨嘴龍及黑亞瓦提龍近緣;但麥當勞2017年的研究則發現最後者較為衍化。[6]
| 鴨嘴龍形類 Hadrosauriformes |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
2018年泰瑞·蓋茨(Terry Gates)等人敘述了與高吻龍來自同一地區的禽龍類新屬喬伊爾齒龍,過去被認為是原賴氏龍的近親。他們的種系發生學分析以諾曼2015年的資料庫為依據,發現喬伊爾齒龍是原賴氏龍的姊妹分類單元。這項分析未採用常見的原賴氏龍+始鴨嘴龍姊妹群。這種親緣關係有三個特徵來支持:一個是梯形枕骨,另外兩個是牙齒的共有衍徵。這兩屬相對於其他鴨嘴龍形類的分類位置取決於,喬伊爾齒龍有無眶前孔,雖然正模標本的幼體有一個孔,但不排除成年後該孔會逐漸閉合的可能性。以下兩個演化樹,分別使用蓋茨等人前述的特徵有無來區分,其中前者形成一個大的多分支:[33]
|
A版本:喬伊爾齒龍眶前孔還在[33]
|
B版本:喬伊爾齒龍的眶前孔後來封閉[33]
|
古生物學
蛋殼碎片連帶單獨的牙齒曾在數個地方發現過,可能屬於原賴氏龍。這些蛋殼表面有網狀紋路,厚度約2毫米。[2]
發育
因為原賴氏龍從幼體到成體都有為數眾多的化石發現,對於了解牠們的成長過程變化提供了充分的證據記錄。上頜骨牙齒數量隨著年齡增長而增加,從幼體的23顆增至成體的33顆。齒骨的牙齒也呈現類似的增長模式,從幼體的18-22顆增至成體的25-30顆。雖然幼體齒骨牙齒的齒冠上缺乏次生脊,但在成體中出現非常細微的次生脊。[3]如科克蘭的研究所述,坐骨腳的靴狀突尺寸也隨年齡增長而增加。[2]坐骨軸的筆直度也可能有所變化,如同FMNH標本顯示出的情況。[6]

2017年麥當勞等人記載了針對FMNH PR 3847標本的其中一根肋骨進行的骨骼組織學分析成果。肋骨內部基質發生了大幅度的骨重塑。骨髓腔(骨髓儲存處)以含有大量骨穴空腔為明顯特徵。在骨骼的外表面上,骨單位幾乎徹底取代了骨頭的內部空間;骨骼分層生長在內部更加明顯,可觀察到五條成長停滯線(lines of arrested growth,LAGs)。髓質區可能有第六條成長停滯線,但範圍相對受到侷限,使得較難鑑定。[6]
因為成長停滯線會在骨重塑過程中被抹除,代表實際上的成長停滯線數目可能被大幅低估。麥當勞等人在檢驗標本時估計在每兩條仍可見的成長停滯線之間的間隙中,至少還有三到四條已消失的成長停滯線。這代表著該動物死亡時只有八到九歲。越接近骨骼外表面,成長停滯線之間的間隙明顯變小,顯示出成長正逐漸趨緩,但外部基本系統(指一系列緊密排列的成長停滯線)的缺乏則代表成長尚未停止。以上情況符合身體其他部位的未成熟跡象,如頸椎、背椎神經棘與椎體之間的縫線尚未癒合,以及薦骨椎體與對應肋骨間的縫線尚未癒合。[34]整體來看,原賴氏龍的成熟速率與亞冠龍、慈母龍差不多,約七至八年。[6][35][36]
古生態學
環境與埋藏學
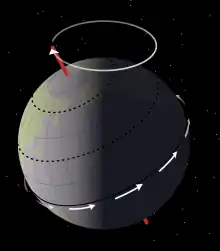
穆森圖奇段的沉積學數據顯示原賴氏龍生活於一個排水不良的氾濫平原,環繞著一系列大型、恆久的湖泊。湖泊水位高低間的波動規律大約落在21000年的米蘭科維奇循環,由地軸自轉的歲差所驅動。這樣的循環模式反映於現在岩石沉積物中的泥質砂岩與泥岩交替層。乾涸時期,裸露的湖床構成一大片無植物生長的寬闊灘地。湖床泥裂產生出碎屑岩,而有機碎屑和屍體殘渣隨著湖水消退散落於水岸邊。與此同時當海侵期間,小河流入氾濫平原,形成生物擾動並汰換掉旱季沉積的碎屑。[5]

穆森圖奇段發現的遺骸大多是關節脫落及四散的,完整骨骼相對罕見。雖然湖泊沒有直接破壞骨頭,但由於埋藏過程的時間拉得特別長,不利於良好保存。在埋藏之前骨骼材料可能已經被清除者破壞,例如西菲力2號採石場發現的原賴氏龍脛骨上有齒痕。湖泊回漲可能偶爾形成沼澤,但持續時間不足以困住及保存較大型脊椎動物。水流作用似乎在西菲力2號採石場的原賴氏龍骨骼沉積過程發揮影響,它們主要沉積的方位(相對北方)為28°、69°、93°、131°、161°。除了69°和93°可能代表河水流向,其餘角度皆大致與湖邊平行,顯示出湖泊流向。[5]
豐富的植物碎屑,加上缺乏內流沉積物,代表在穆森圖奇段沉積期間,當地屬於相對潮濕的氣候環境。與當地較古老的沉積物形成明顯的對比,後者呈石灰質,其厚度顯示乾旱期長達10000年之久。古氣候重建支持這些理論:莫里海(Mowry Sea)自北方滲入(最終將形成西部內陸海道),造成原本偏乾燥的溫帶大陸型氣候,逐漸被潮濕的溫帶海洋氣候所取代。[37]根據全球氣候模式顯示,穆森圖奇的古緯度為46°N,表面均溫為40-45°C,比現代暖約6-8°C。[5][38]
生態系

穆森圖奇沉積層盛產鱷形超目,以發現牙齒為主。已知最大的牙齒被歸入大頭鱷科;[lower-alpha 1]牙齒寬闊、基部呈圓形,某些帶有縱脊。[6]這些大型鱷類至少有能力掠食岸邊的原賴氏龍幼體。發現的湖邊化石包含大量幼體標本支持此理論;以鱷類來說無法對體型較大的成體造成威脅。[5]此外亦發現較小顆的鱷類牙齒,呈現出兩種不同形態:帶脊的卵圓形被歸入伯尼斯鱷的兩個物種;[5][lower-alpha 1]收縮三角形帶有厚紋飾被歸入阿吐波鱷科。[11][40][lower-alpha 1]然而有鑑於這些牙齒平均尺寸偏小,且某些鱷類同時存在著多種形態的牙齒,[39]實際上可能都屬於同一物種。[6]

同樣由菲爾德博物館團隊發掘的[6]新獵龍科西雅茨龍[41]可能對較大隻的原賴氏龍造成威脅。[5]穆森圖奇段還有數種較小型的手盜龍類,包含馳龍科、近爪牙龍、理察伊斯特斯龍,牠們可能佔據著食腐動物的棲位。[40]至於植食性恐龍方面,有出土於卡蘿點的結節龍科活堡龍,位置接近原賴氏龍的模式標本。[42]還發現更多屬於基礎鳥腳類的牙齒,曾由西菲力於1999年提及為厚頭龍類或有齒鳥類。[5][40]此外來自兩種不同鳥腳類:一種奔山龍亞科和一種基礎禽龍類的更完整遺骸至今尚未發表。[43]一種巨型偷蛋龍類也與屬於角龍類和蜥腳類的牙齒一同被發現。[40][44][6]
較小型的鱷類可能食用魚類,牠們的發現以鱗片和牙齒為主,包含燕魟目的Baibisha;[40]一種未定新鰭亞綱;旗背魚類的某種鱗齒魚;一種與福鱷有關的雀鱔科;[6]堅齒魚科的Stephanodus;弓鰭魚目未定種。亦有發現未定有尾目(蠑螈)和哺乳類。至於植物方面,花粉標本顯示穆森圖奇湖泊周遭分布著一片層狀森林,以桫欏目、南洋杉屬、松柏門構成林冠;下層長著蕨類、裸子植物、被子植物。湖泊等水域也會出現藻類。[8][45]這些植物為當地的植食性恐龍如原賴氏龍提供了充沛的食物來源。[5]
古生物地理學
科克蘭、西菲力等人指出,穆森圖奇段的動物相(禽龍類、厚頭龍類、角龍類)與同時期的亞洲動物相有非常高的相似程度。他們表示原賴氏龍屬於森諾曼階亞洲恐龍遷徙湧入北美的一個實例,這些外來種取代了早期多樣性低落的原生種動物相。[2][11]原賴氏龍與原巴克龍(黑德的種系發育分析)、福井龍(蘇伊士及阿維葉諾夫的種系發育分析)的近緣關係支持此理論。[3][14][27]
然而霍納等人、皮耶妥·馬奎茲、麥當勞等人的研究結果則與此理論矛盾,他們認為同樣來自北美的始鴨嘴龍才與原賴氏龍近緣。[6][26][28][46]這樣不同的觀點顯示原賴氏龍、始鴨嘴龍、乃至更衍化的鴨嘴龍科的共同祖先不僅僅來自亞洲,而是同時分布於亞洲和北美洲。[3]麥當勞等人進一步指出,原賴氏龍和始鴨嘴龍代表侷限於北美的特有種鴨嘴龍類,如同白堊紀其他地方的直拇指龍類族群,例如同時代的中亞特有種包含雙廟龍、張衡龍、近鴨嘴龍。[6]
參考來源
| 维基共享资源上的相关多媒体资源:原賴氏龍屬 |
- Galton, Peter; Jensen, James A. (PDF). Brigham Young University Geology Studies. 1979, 25 (3): 1–10 [2021-05-27]. ISSN 0068-1016. (原始内容存档 (PDF)于2021-01-26).
- Kirkland, J.I. . Lucas, S.G.; Kirkland, J.I.; Estep, J.W. (编). . New Mexico Museum of Natural History and Science Bulletin 14. 1998: 283–295.
- McDonald, A.T.; Bird, J.; Kirkland, J.I.; Dodson, P. . PLOS ONE. 2012, 7 (10): e45712. Bibcode:2012PLoSO...745712M. PMC 3471925
 . PMID 23077495. doi:10.1371/journal.pone.0045712.
. PMID 23077495. doi:10.1371/journal.pone.0045712. - Kirkland, J.I.; Burge, D. . . Fifty-Fourth Annual Meeting, Society of Vertebrate Paleontology 14 (3): 32A. 1994. JSTOR i406457.
- Garrison, J.R. Jr.; Brinkman, D.; Nichols, D.J.; Layer, P.; Burge, D.; Thayn, D. . Cretaceous Research. 2007, 28 (3): 461–494. doi:10.1016/j.cretres.2006.07.007.
- McDonald, A.T.; Gates, T.A.; Zanno, L.E.; Makovicky, P.J. . PLOS ONE. 2017, 12 (5): e0176896. Bibcode:2017PLoSO..1276896M. PMC 5425030 . PMID 28489871. doi:10.1371/journal.pone.0176896.
- Craig, L.C. . Rocky Mountain Association of Geologists 1981 Field Conference. 1981: 195–200 [2021-05-30]. (原始内容存档于2021-06-03).
- Tschudy, R.H.; Tschudy, B.D.; Craig, L.C. (报告). US Geological Survey, Professional Paper 1821. US Geological Survey: 1–24. 1984 [2021-05-30]. (原始内容存档于2021-06-03).
- Nichols, D.J.; Sweet, A.R. . Caldwell, W.G.E.; Kauffman, E.G. (编). . Geological Association of Canada, Special Paper 39. Geological Association of Canada. 1993: 539–584.
- Young, R.G. . American Association of Petroleum Geologists Bulletin. 1960, 44 (2): 156–194 [2021-05-30]. doi:10.1306/0bda5fd2-16bd-11d7-8645000102c1865d. (原始内容存档于2021-06-03).
- Cifelli, R.L.; Kirkland, J.I.; Weil, A.; Deino, A.L.; Kowallis, B.J. . Proceedings of the National Academy of Sciences. 1997, 94 (21): 11163–11167. Bibcode:1997PNAS...9411163C. PMC 23402 . PMID 9326579. doi:10.1073/pnas.94.21.11163.
- Paul, G.S. . . Princeton: Princeton University Press. 2010: 327 [2021-05-27]. ISBN 9780691167664. (原始内容存档于2019-04-08).
- Holtz, T.R. (PDF). Supplementary Information to Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages by Thomas R. Holtz, Jr., illustrations by Luis Rey. Department of Geology, University of Maryland. 2012 [2017-08-30]. (原始内容存档 (PDF)于2017-08-12).
- Head, J.J. . Journal of Vertebrate Paleontology. 2001, 21 (2): 392–396. doi:10.1671/0272-4634(2001)021[0392:AROTPP]2.0.CO;2.
- Weishampel, D.B.; Norman, D.B.; Grigorescu, D. . Palaeontology. 1993, 36 (2): 361–385 [2021-07-24]. (原始内容存档于2021-07-25).
- Sereno, P.C. . National Geographic Research. 1986, 2: 234–256.
- Brett-Surman, M.K. (PDF) (学位论文). Washington D.C.: George Washington University: 1–373. 1989 [2021-07-24]. (原始内容存档 (PDF)于2017-09-10).
- Weishampel, D.B.; Horner, J.R. . Weishampel, D.B.; Dodson, P.; Osmolska, H. (编). 1st. Berkeley: University of California Press. 1990: 534–561.
- Horner, J.R.; Currie, P.J. . Carpenter, K.; Hirsch, K.F.; Horner, J.R. (编). . Cambridge: Cambridge University Press. 1994: 312–336 [2021-07-24]. ISBN 9780521567237. (原始内容存档于2021-07-24).
- Casanovas, M.L.; Pereda Suberbiola, X.; Santafe, J.V.; Weishampel, D.B. . Geological Magazine. 1999, 136 (2): 205–211. Bibcode:1999GeoM..136..205C. doi:10.1017/s0016756899002319.
- Lull, R.S.; Wright, N.E. . Geological Society of America Special Papers. 1942, 40: 1–242. doi:10.1130/SPE40-p1.
- Norman, D.B. . Bulletin de l'Institut Royal des Sciences Naturelles de Belgique. 1986, 56: 281–372 [2021-07-24]. (原始内容存档于2017-09-10).
- Norman, D.B. . Zoological Journal of the Linnean Society. 1998, 122 (1): 291–348. doi:10.1111/j.1096-3642.1998.tb02533.x.
- Norman, D.B. . Weishampel, D.B.; Dodson, P.; Osmólska, H. (编). 2nd. Berkeley: University of California Press. 2004: 413–437 [2021-07-25]. (原始内容存档于2021-07-25).
- Head, J.J. . Journal of Vertebrate Paleontology. 1998, 18 (4): 718–738. doi:10.1080/02724634.1998.10011101.
- Horner, J.R.; Weishampel, D.B.; Forster, C.A. . Weishampel, D.B.; Dodson, P.; Osmólska, H. (编). 2nd. Berkeley: University of California Press. 2004: 438–463 [2021-07-25]. (原始内容存档于2021-07-25).
- Sues, H.-D.; Averianov, A. . Proceedings of the Royal Society B. 2009, 276 (1667): 2549–2555. PMC 2686654 . PMID 19386651. doi:10.1098/rspb.2009.0229.
- Prieto-Márquez, A.; Norell, M.A. . American Museum Novitates. 2010, 3694: 1–49. S2CID 56372891. doi:10.1206/3694.2.
- Zheng, W.; Jin, X.; Shibata, M.; Azuma, Y. . Historical Biology. 2014, 26 (2): 236–251. S2CID 129081459. doi:10.1080/08912963.2013.809347.
- Gasca, J.M.; Moreno-Azanza, M.; Ruiz-Omeñaca, J.I.; Canudo, J.I. . Journal of Iberian Geology. 2015, 41 (1): 57–70. doi:10.5209/rev_jige.2015.v41.n1.48655 .
- Verdú, F.J.; Royo-Torres, R.; Cobos, A.; Alcalá, L. . Cretaceous Research. 2015, 56: 250–264. doi:10.1016/j.cretres.2015.05.010.
- Verdú, F.J.; Royo-Torres, R.; Cobos, A.; Alcalá, L. . Historical Biology. 2017, 30 (4): 437–474. S2CID 89715643. doi:10.1080/08912963.2017.1287179.
- Gates, T.A.; Tsogtbaatar, K.; Zano, L.E.; Chinzorig, T.; Watabe, M. . PeerJ. 2018, 6: e5300. PMC 6078070 . PMID 30083450. doi:10.7717/peerj.5300.
- Brochu, C.A. . Journal of Vertebrate Paleontology. 1994, 16 (1): 49–62. doi:10.1080/02724634.1996.10011283.
- Horner, J.R.; de Ricqlès, A.; Padian, K. . Paleobiology. 1999, 25 (3): 295–304. doi:10.1017/S0094837300021308.
- Woodward, H.N.; Freedman Fowler, E.A.; Farlow, J.O.; Horner, J.R. . Paleobiology. 2015, 41 (4): 503–527. S2CID 85902880. doi:10.1017/pab.2015.19.
- Elder, W.P.; Kirkland, J.I. . Caputo, M.V.; Peterson, J.A.; Franczyk, K.J. (编). . Denver: Rocky Mountain Section SEPM. : 415–440 [2021-08-04]. (原始内容存档于2021-08-04).
- Glancy, T.J. Jr.; Arthur, M.A.; Barron, E.J.; Kaufman, E.G. . Caldwell, W.G.E.; Kauffman, E.G. (编). . Geological Association of Canada, Special Papers 39. Geological Association of Canada. 1993: 219–241.
- Irmis, R.B.; Hutchison, J.H.; Sertich, J.J.W.; Titus, A.L. . Titus, A.L.; Loewen, M.A. (编). . Bloomington: Indiana University Press. 2013: 424–444 [2021-08-09]. ISBN 9780253008961. (原始内容存档于2021-08-09).
- Cifelli, R.L.; Nydam, R.L.; Gardner, J.D.; Weil, A.; Eaton, J.G.; Kirkland, J.I.; Madsen, S.K. . Gillette, D. (编). . Utah Geological Survey Miscellaneous Publications 99–1. 1999: 219–242.
- Zanno, L.E.; Makovicky, P.J. . Nature Communications. 2013, 4: 2827. Bibcode:2013NatCo...4.2827Z. PMID 24264527. doi:10.1038/ncomms3827 .
- Carpenter, K.; Kirkland, J.I.; Burge, D.L.; Bird, J. . Gillette, D. (编). . Utah Geological Survey Miscellaneous Publications 99–1. 1999: 243–251 [2021-08-09]. (原始内容存档于2021-08-09).
- Zanno, L.E.; Makovicky, P.J. . Society of Vertebrate Paleontology 76th Annual Meeting. Salt Lake City: Society of Vertebrate Paleontology: 256. 2016. (原始内容使用
|archiveurl=需要含有|url=(帮助)存档于2016-10-18). - Makovicky, P.J.; Zanno, L.E. Kirkland, J.I.; Hunt-Foster, R.; Liggett, G.A.; Trujillo, K. , 编. . Mid-Mesozoic: The Age of Dinosaurs in Transition (PDF). Fruita: Utah Geological Survey: 70–71. 2014. (原始内容存档 (PDF)于2014-10-06) 使用
|archiveurl=需要含有|url=(帮助). - Ravn, R.L. . Palaeontographica Abteilung B. 1995, 234 (3): 41–91 [2021-08-09]. (原始内容存档于2021-08-09).
- Prieto-Márquez, A. . Zoological Journal of the Linnean Society. 2010, 159 (2): 503–525. doi:10.1111/j.1096-3642.2010.00642.x .